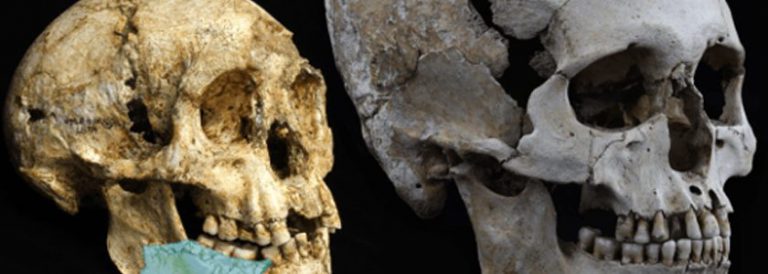
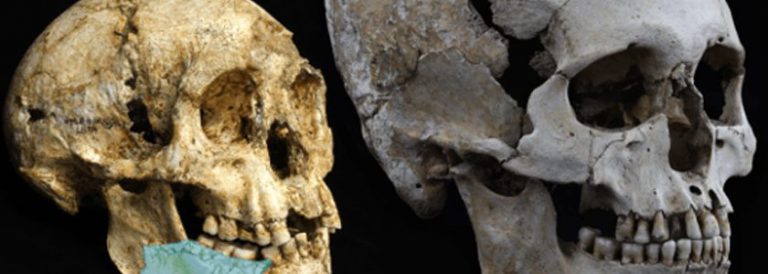
L’evolució del cervell (2). (Som les nostres connexions 12)
6 juliol, 2016
«Desafortunats els científics que només tinguin en el seu cap idees clares»
Louis Pasteur
Reflexió inicial
Potser les preguntes més suggeridores que la ciència actual té plantejades són l’origen de l’Univers, l’emergència de la vida i l’aparició de la nostra espècie. Aquests tres orígens van suposar punts d’inflexió respecte de la situació en què es produïen, i van desencadenar esdeveniments d’emergència i innovació transcendentals en aquest llarg camí transcorregut des del Big Bang fins als nostres dies. Així, l’origen de l’Univers va comportar l’aparició del temps i de l’espai, de l’energia i de la matèria. Per la seva banda, l’origen de la vida va suposar l’emergència d’un tipus especial d’entitats químiques que tenen la capacitat de reproduir-se i evolucionar per selecció natural. El tercer origen va ser el que va permetre, dins el llinatge dels primats, l’aparició del gènere Homo: una successió d’espècies caracteritzades per un progressiu desenvolupament del seu cervell, associat a l’ús cada vegada més perfeccionat d’eines. Així es va arribar als humans moderns, a nosaltres: els únics éssers vius coneguts que som capaços de preguntar-nos pel nostre lloc en el cosmos.
Són nombrosos els interrogants que es plantegen, molts dels quals són la versió actualitzada de les preguntes que ens han acompanyat des de sempre : qui som?, d’on venim?, estem sols a l’Univers?, té sentit tot plegat? quina funció té la consciència? per què i per a què ens podem formular aquestes preguntes?
El que és impossible ara també ho va ser en el passat, ja que les regles fisicoquímiques del joc no han canviat des de la primera fracció de segon de l’Univers. Dins del territori del que és possible, sabem que hi ha fenòmens amb més o menys probabilitat d’haver-se produït: a partir dels més probables hem de ser capaços de construir un relat lògic que ens permeti avançar cap al que va poder ocórrer en cadascun dels orígens.
I aquest relat ha de començar exposant les possibles raons que, dins de la branca dels primats –originats fa entre 74 i 63 milions d’anys–, van produir el «big bang» del llinatge del gènere Homo: què és el que fa humà a l’ésser humà? Quins caràcters ens defineixen com a espècie? Si ens comparem amb els nostres més propers parents, els ximpanzés, aquesta pregunta sembla fàcil de respondre, malgrat compartir més del 90 per cent del nostre genoma.
- el nostre cervell és gairebé quatre vegades més gran, amb un volum d’uns 1.400 cc, enfront dels 300 o 400 cc del d’un ximpanzé.
- el nostre tipus de locomoció és també molt diferent. Nosaltres practiquem una marxa bípeda que difereix en gran mesura del tipus de locomoció que practiquen goril·les i ximpanzés. Aquests últims s’ajuden amb els artells de les mans per desplaçar-se sobre el sòl (l’anomenat knuckle-walking), en un tipus de locomoció que podríem qualificar de semiquadrúpeda. Tot i que els ximpanzés pigmeus o bonobos poden desenvolupar també una marxa bípeda durant algun temps, nosaltres no som bípedes ocasionals, sinó obligats per la nostra pròpia anatomia, com també ho són, per exemple, les aus. El nostre aparell locomotor està adaptat per a desenvolupar una marxa forçosament bípeda i qualsevol altre tipus de locomoció ens resulta desmanegat quan no impossible. Això es manifesta, per exemple, en la diferent configuració dels nostres peus i mans. Mentre que les dues extremitats presenten en ximpanzés i goril·les una anatomia semblant, en nosaltres el peu s’ha adaptat eficaçment a la locomoció bípeda, de manera que ha perdut la seva capacitat de prensió .
- no tenim la gruixuda capa de pèl que recobreix la pell dels nostres parents més propers. El fet de ser «micos nus» [1], ens priva d’algunes de les tècniques d’apaivagament i estructuració social que són característiques dels ximpanzés, com en el cas de l’esplugament mutu.
- per a comunicar-nos fem servir un llenguatge complex, basat en fonemes que es relacionen entre ells segons una estructura sintàctica.
- som capaços de manufacturar tot tipus d’estris, alguns dels quals tenen una evident utilitat pràctica, però molts altres corresponen a objectes la funció dels quals és molt més simbòlica que utilitària, com és el cas de joies, collarets…
- som capaços d’elaborar representacions més o menys simbòliques del món que ens envolta. En altres paraules, som animals culturals.
- tenim una capacitat inusitada per transmetre aquesta cultura a altres membres del grup mitjançant l’aprenentatge.
No és que característiques com la comunicació, la cultura o l’aprenentatge no es donin també en altres espècies, però només en nosaltres apareixen associades entre si i portades fins a extrems de complexitat que no trobem en els nostres parents més propers.
Segons les dades moleculars i paleontològics, el nostre últim avantpassat comú amb els ximpanzés va viure a l’Àfrica fa uns 7 o 8 milions d’anys i gràcies a les evidències paleontològiques i arqueològiques, podem rastrejar com el nostre llinatge ha anat acumulant el conjunt de característiques que avui ens permeten reconèixer-nos com una espècie singular.
Origen, evolució i classificació dels primats
Els humans actuals formem part de la diversitat dels éssers vius i només nosaltres hem arribat a ser capaços de fer-nos preguntes sobre l’origen de l’Univers, l’origen de la vida i sobre nosaltres mateixos.
Pertanyem a l’ordre Primats (en llatí, «els primers»). El 1758 el naturalista suec Carl von Linné (1707-1778) va batejar amb aquest nom a totes les espècies de mamífers semblants a nosaltres, seguint la tradició antropocèntrica encara imperant en el seu temps. Es va fixar en les similituds que ens uneixen a les espècies vives de primats i ens va classificar amb ells. Linné no va ser capaç de trobar evidències orgàniques per situar-nos en algun grup diferent i privilegiat, com hauria estat del gust de la societat de l’època.
El seu origen es situa entre 74 i 63 milions d’anys, al límit entre el Cretaci (145 – 66 milions) i el Cenozoic (66 milions – actualitat) –alguns estudis moleculars arriben fins als 85 milions.
Aspecte dels primers primats: els mamífers d’aquella remota època més propers a la nostra genealogia eren els avantpassats dels rosegadors i dels lagomorfs (conills i llebres).
Alguns investigadors han assignat les primeres restes fòssils conegudes d’un primat al gènere Plesiadapis (altres científics aposten per espècimens fòssils atribuïts, entre d’altres, als gèneres Teilhardina, Altanius, Cantius o Archicebus). La seva reconstrucció ens recorda un esquirol estilitzat, de potes llargues i amb els ulls encara situats en posició lateral. De seguida arribaria la migració dels ulls cap a una posició frontal i la visió binocular, gairebé sempre en color.
Durant la seva evolució, els primats han mantingut caràcters primitius, com la presència de cinc dits i una dentició poc especialitzada, generalment de dents proveïts de cúspides arrodonides, aptes per al consum d’una dieta omnívora. Van evolucionar cap a la pèrdua d’urpes en els dits i la seva substitució per ungles planes, la disminució del musell i l’increment del cervell, en particular del còrtex cerebral.
La classificació dels primats ha anat variant al llarg dels anys. La millora en les observacions de les espècies vives i dels fòssils, així com l’estudi del DNA, ha contribuït a millorar aquesta classificació. En l’actualitat, els primats inclouen dos subordres: estrepsirins (del grec strepho, corb i rhis, nas) i haplorins (del grec haplos, simple i rhinos, nas). Segons ens revelen les anàlisis moleculars, aquests dos grups van divergir gairebé des dels inicis de la història evolutiva dels primats.
Esquelet de Darwinius masillae de 47 milions d’anys d’edat, és un dels primats fòssils més complets mai trobats. Prové de la localitat de Messel, prop de la ciutat de Darmstadt, a Alemanya. Conserva tot l’esquelet, excepte la cama esquerra per sota del genoll.
Varietat de primats estrepsirins
Els estrepsirins es caracteritzen pel seu musell humit, que connecta amb la boca i està recobert per una membrana (el rinari). Aquest subordre inclou els lorísids, lemúrids i gàlags.
Varietat de primats haplorins
Els haplorins no tenen rinari, el seu nas no està connectat amb la boca i tampoc posseeixen virbrisses (pèls especialitzats a banda i banda del musell, utilitzats per al sentit del tacte). Es van dividir, fa uns 58 Ma, en dos infraordres[2]: Tarsiiformes i Simiiformes.
Els Simiiformes ens conduiran cap al nostre origen. Aquest grup va divergir fa uns 40 milions d’anys, quan part d’ells (Platyrrhini) va colonitzar Amèrica del Sud des d’Àfrica. El grup dels platirins (micos del Nou Món), són primats de talla petita, cues llargues i prènsils i musells aplatats, amb narius (fosses nasals) en posició lateral. L’altre grup (Catarrhini) es va quedar a l’Àfrica i va colonitzar Euràsia en els inicis del Miocè, fa uns 23 milions d’anys. Les narius dels catarins s’obren cap avall i estan separades per un envà nasal. Aquest grup es coneix com els micos del Vell Món i els seus representants vius estan inclosos en les famílies dels cercopitècids, hilobàtids i homínids. En aquesta última (Hominidae) ja ens trobem amb els orangutans, goril·les i ximpanzés, els nostres parents més propers tant des del punt de vista del temps geològic com de les diferències entre els nostres respectius genomes.
A la recerca de l’avantpassat comú
La família Hominidae (homínids) inclou quatre gèneres amb representants actuals: Gorilla, Homo, Pan i Pongo. El genoma de les set espècies reconegudes en aquests quatre gèneres presenta una gran similitud, evidenciant una divergència relativament recent entre elles. Els humans actuals compartim un antecessor comú amb el llinatge dels ximpanzés, els simis més propers a nosaltres en termes evolutius.
L’evolució humana es podria descriure com un llibre al que li falten el 98% de les seves pàgines i del qual no en tenim el pròleg i dels primers capítols just en queden algunes frases inconnexes. No sabem com comença la història i hem de fer un gran esforç d’imaginació per reconstruir l’escenari evolutiu, així com els intèrprets que van representar el primer acte de la història de la genealogia humana.
En comparar el genoma d’Homo sapiens i el de Pan troglodytes (una de les espècies de ximpanzé) s’infereix que compartim un antecessor comú, que va viure a l’Àfrica a finals del Miocè fa entre 7 i 6 Ma. Abans del 8 Ma, la major part d’Àfrica estava coberta de boscos i només després va començar una progressiva desforestació de certes regions. A finals del Miocè (23,03 – 5,3 milions) l’est del continent va començar a refredar-se i va canviar el seu paisatge per efecte de l’elevació de la Gran Vall del Rift.
L’antecessor comú d’humans i ximpanzés va haver de viure en l’ambient boscós d’algun recòndit lloc del continent africà. La presència de selves a la regions més càlides i humides d’Àfrica va evitar la conservació de les evidències fòssils que es necessiten per contrastar qualsevol hipòtesi sobre el que va succeir en aquesta època: quin aspecte tenia l’antecessor comú?, com era la seva biologia?, quina era la seva forma de locomoció?
El tema de la locomoció és fonamental, ja que la primera i gran diferència entre els membres de les dues genealogies és la manera en què uns i altres ens movem en els nostres desplaçaments. Els ximpanzés són quadrúpedes, encara que siguin capaços de mantenir-se drets durant breus lapses de temps. L’arquitectura del seu esquelet està preparada per enfilar-se amb facilitat, però utilitzen les quatre extremitats quan volen avançar per terra a gran velocitat. L’ús dels artells durant la locomoció és característic dels ximpanzés i dels goril·les. Els membres de la genealogia humana caminem drets i som bípedes.
No sabem com es desplaçava el nostre avantpassat comú. No en tenim ni una sola evidència. Els medis humits i boscosos reciclen la matèria orgànica amb extrema rapidesa i en aquestes regions amb prou feines hi ha llocs proclius a la fossilització dels éssers vius.
Tant de bo es trobés un esquelet del nostre avantpassat comú amb els ximpanzés i, en particular, la seva pelvis. Podríem entendre llavors com es va produir la transició entre la marxa quadrúpeda i la marxa bípeda. No podem assumir sense més que el nostre avantpassat comú es desplaçava igual que els ximpanzés, però tampoc podem assegurar que fos bípede. En aquest últim cas, els ximpanzés haurien perdut la facultat de caminar només amb les extremitats inferiors en el transcurs de la seva evolució.
1. Els primers homínids bípedes
Durant bona part del segle XX, la investigació sobre l’evolució humana va estar condicionada per la idea que la principal característica que definia al nostre llinatge era la possessió d’un gran cervell. Aquest mateix prejudici va impedir que durant anys la comunitat científica acceptés com homínid a Australopithecus africanus, l’espècie erigida pel metge sud-africà Raymond Dart el 1925 a partir d’un crani infantil, malgrat que el crani mostrava alguns trets inequívocament humans:
- posició avançada del foramen magnum (indicador d’una posició bípeda)
- canines de mida reduïda,
el seu cervell era comparable al d’un ximpanzé i, per tant, la seva naturalesa homínida qüestionable.
Amb l’aparició de noves restes esquelètiques i no només cranials d’Australopithecus, la idea que els nostres avantpassats estaven dotats d’un cervell petit i, alhora, eren bípedes, es va anar obrint pas, i es va confirmar totalment amb la troballa, el 1974, per Donald E. Johanson i Tim White, de la «Lucy», una Australopithecus afarensis, un homínid perfectament bípede.
Esquema de la meitat inferior del cos humà mostrant la funció dels músculs abductors durant la locomoció bípeda.
Articulació del fèmur amb la tíbia en Homo sapiens (a), Australopithecus afarensis (b) i en un ximpanzé (c).
Les restes de Lucy se situaven en dipòsits datats en uns 3,5 milions d’anys. Ara bé, ¿fins quan es pot retrotraure l’origen de la postura bípeda?
Sahelanthropus tchadiensis
Sabem que el nostre llinatge es va escindir del dels ximpanzés fa uns 7 milions d’anys. Seria, per tant, al voltant d’aquesta data quan es podrien trobar els primers homínids bípedes. Les restes més antigues atribuïbles a un homínid bípede són les de la localitat de Toros-Menalla 266, al Txad. Les troballes publicades per Michel Brunet[3] i el seu equip l’any 2002 corresponen a un crani deformat però bastant complet i que, juntament amb algunes peces dentàries, van servir per definir un nou gènere i espècie, Sahelanthropus tchadiensis.
- l’edat atribuïda a aquesta localitat se situa precisament entre 6 i 7 milions d’anys just en el moment predit per la biologia molecular per trobar els primers homínids bípedes.
- el crani amb una escassa capacitat craniana (entre 320 i 380 cc comparable a la d’un ximpanzé), presenta una combinació de trets que l’acosten tant als homínids posteriors com als actuals antropomorfs africans.
- existència d’una prominent cresta o torus supraorbital a manera de visera, que recorda a la dels goril·les.
- comú també als goril·les és la presència d’una cresta sagital a la part posterior del crani.
- per sota d’unes òrbites més o menys arrodonides la cara és curta i plana poc projectada cap endavant, a diferència del que succeeix en els antropomorfs africans actuals en què la cara és prognàtica.
- la dentició presenta esmalt prim (encara que més gruixut que en els actuals ximpanzés) i és relativament petita. Les dents estan implantats verticalment sobre el maxil·lar mentre que les canines són petites i fortament desgastades.
- – un element clau per saber si Sahelanthropus tchadiensis era bípede és la posició del foramen magnum (l’orifici cranial que allotja el principi de la columna vertebral); planteja dubtes atès que es tracta d’un crani deformat.
Orrorin tugenensis
Posteriors en edat són les restes recuperades al jaciment kenyà de Lukeino, datats en uns 6 milions d’anys. Aquestes restes, corresponents a tres fragments mandibulars, diverses peces dentàries superiors, dos fèmurs incomplets, un fragment d’húmer, van permetre a l’equip dirigit per Brigitte Senut i Martín Pickford, del Museu Nacional d’Història Natural de París reconèixer un nou tipus d’homínid, al que van batejar amb el nom de Orrorin tugenensis.
- la forma del cap del fèmur, esfèrica, indica que Orrorin tugenensis mostrava ja una locomoció bípeda. Però és qüestionat perquè les proves no són prou convincents.
- la presència de falanges manuals recorbades o la forma de l’húmer, indicaven que Orrorin mantenia encara una bona capacitat per enfilar-se pels arbres.
- les dents d’Orrorin eren relativament petites en comparació amb els d’Australopithecus, però tenien en comú amb aquest últim la presència d’esmalt dentari gruixut. Les canines eren comparativament grans, en relació amb els d’altres homínids.
Ardipithecus ramidus
Molt més completa que els anteriors és la mostra procedent del jaciment d’Aramis, a la regió de l’Awash mitjà, a Etiòpia (White et al., 2010). Aquest jaciment, datat en uns 4,4 milions d’anys és bastant més recent que els anteriors, però encara més antic que la més antiga evidència d’Australopithecus. El jaciment d’Aramis ha proporcionat nombroses restes (fins a 36 individus) de l’espècie Ardipithecus ramidus, entre ells l’esquelet parcialment complet d’una femella jove.
- fou un petit homínid de no més de 50 quilos de pes i una alçada d’uns 120 cm.
- el seu cervell tot just sobrepassava els 300 grams i era, per tant, similar en grandària al d’un ximpanzé.
- la dentició presentava un esmalt prim, com en goril·les i ximpanzés. A diferència d’aquests, però, les canines eren de mida reduïda, no només en les femelles sinó també en els mascles.
- pel que fa al seu aparell locomotor, les mans mostraven unes falanges llargues i corbades, el que indica un tipus de locomoció encara associada a la vida en els arbres.
- els peus presentaven un dit gros separat de la resta de dits i amb capacitat prènsil, com succeeix en ximpanzés i goril·les.
- els braços eren relativament llargs, la qual cosa indica una bona capacitat per desplaçar-se de branca a branca.
- al costat d’aquest cúmul de característiques lligades al bosc, algunes dades semblen donar suport que Ardipithecus era ja un homínid bípede capaç de desplaçar-se a terra sobre els seus dos peus. La part superior de la pelvis presenta un aspecte més semblant a la dels homínids posteriors que a la de ximpanzés i goril·les. El foramen magnum, així mateix, presenta una posició més avançada que en aquests últims, suggerint que la columna vertebral se situava ja per sota del crani i no a continuació d’ell.
Ardipithecus kadabba
El registre fòssil d’Ardipithecus s’ha pogut estendre gairebé un milió d’anys enrere gràcies al descobriment d’una segona espècie, A. kadabba, en nivells datats entre 5,2 i 5,8 milions d’anys de la regió del Middle Awash, a Etiòpia.
– els autors de la troballa argumenten, basant-se algunes restes del peu, que Ardipithecus kadabba era ja un homínid bípede.
Australopithecus anamensis
La primera evidència d’un homínid incontrovertiblement bípede procedeix dels jaciments de Kanapoi i Allia Bay, a la vora del llac Turkana, a Kenya. Es tracta d’Australopithecus anamensis, datat en uns 4 milions d’anys i la més antiga espècie reconeixible del gènere Australopithecus.
Restes fòsils de l’ Australopithecus anamensis
- encara que fins al moment no s’han trobat restes de la caixa craniana d’aquesta espècie, sabem que era ja una forma bípeda pel disseny de la seva tíbia, eixamplada en el seu extrem anterior i molt similar a les del nostre propi gènere. Aquest eixamplament de la part superior de la tíbia és característic de tots els homínids bípedes, les extremitats posteriors han de sostenir tot el pes del cos. Per contra, atès que ximpanzés i goril·les practiquen un tipus de locomoció semiquadrúpeda sobre els artells, la part superior de les seves tíbies és més prima i dibuixa una característica silueta en forma de «T».
- les dents d’Australopithecus anamensis estaven dotades d’un esmalt gruixut, com és comú a totes les espècies d’Australopithecus.
- encara manté alguns trets arcaics. Així, les dues branques de les mandíbules discorren en paral·lel, tal com s’observa en les mandíbules dels antropomorfs actuals. En homínids posteriors, en escurçar-se la cara, les mandíbules tendeixen a esbossar un disseny més en «V» que en «U», amb les dues branques divergint a partir de les canines. Per contra, aquest és un caràcter que vincula Australopithecus anamensis amb goril·les i ximpanzés.
2. Per a què serveix el bipedisme?
Després d’un procés que probablement es va iniciar fa uns 6 o 7 milions d’anys, fa una mica menys de 5 milions d’anys trobem a l’Àfrica homínids plenament bípedes el tipus de locomoció dels quals era molt semblant al nostre i diferent del de ximpanzés i goril·les. Ara bé, quines pressions de selecció van motivar aquest canvi? Per què en un moment donat un grup d’homínids va desenvolupar aquesta nova adaptació locomotora? Quins avantatges evolutius va poder aportar la marxa bípeda?
- Una primera conseqüència té a veure amb la mida del cervell. En la mesura que el crani se situa per sobre de la columna vertebral i és sostingut per ella, s’obre la possibilitat teòrica d’un augment del volum d’aquest òrgan sense les constriccions que imposa en un vertebrat quadrúpede el fet de desplaçar el centre de gravetat cap endavant en augmentar el pes del cap. D’aquesta manera, el bipedisme ha estat un dels primers factors que ha permès als humans desenvolupar un cervell de prop de 1.500 grams.
- Una altra conseqüència important del bipedisme va ser l’alliberament de les mans, que van adquirir així una funció bàsicament manipuladora i no locomotora, com encara succeeix entre els antropomorfs. Amb el temps, l’alliberament de les mans va permetre el desenvolupament de la pinça de precisió entre el polze i la resta dels dits, així com la capacitat per elaborar objectes complexos, com va ser el cas de les primeres eines lítiques i la resta dels desenvolupaments culturals posteriors.
L’augment de la mida del cervell o l’alliberament de les mans són conseqüències positives del bipedisme, però probablement no la causa que aquest es seleccionés en el curs de l’evolució humana. En termes adaptatius, serien el que es coneix com exaptacions, és a dir, conseqüències derivades de l’adopció d’una locomoció bípeda, però que en principi no van ser seleccionades directament per a les funcions que després desenvoluparien. Així doncs, podem seguir preguntant-nos per les causes que van portar a l’adopció d’una locomoció bípeda per part d’un grup d’homínids africans.
Per què som bípedes?
1. Comparació de la biomecànica de la marxa quadrúpeda dels simis i del bipedisme dels humans.
Quan comparem la forma de la pelvis (on resideixen els secrets que expliquen la manera de desplaçar-nos) d’un ximpanzé o d’un goril·la amb la d’Homo sapiens, ens sembla impossible que hàgim fet un salt anatòmic tan impressionant sense passar per situacions intermèdies. O camines com ho fa un goril·la i un ximpanzé, o ho fas com nosaltres. Altres formes de moure’s, intermèdies entre la d’uns i altres, semblen poc o gens eficaces, quan no impossibles.
La genètica ens explica que no calen molts canvis (mutacions o reordenaments genètics) en el DNA per aconseguir un resultat cridaner. Una única mutació pot alterar el programa i produir un resultat final molt diferent. Potser el bipedisme va ser conseqüència de canvis en determinats gens reguladors.
Quin són els canvis que considerem necessaris per passar d’un primat quadrúpede a un primat bípede?
L’ili de les pelvis dels simis és llarg i estret, amb una ala ilíaca gairebé plana. Per contra, el nostre ili és curt, molt ample i fortament corbat. Què es va aconseguir amb aquest canvi? Els tres músculs que impulsen els ximpanzés cap endavant (adductors) són el gluti major, gluti mitjà i gluti menor, que s’insereixen en l’ala ilíaca i tenen el seu origen en diferents regions del fèmur. Els tres músculs actuen a l’uníson quan aquests animals caminen o corren a tota velocitat, però són incapaços de sostenir als nostres parents quan aquests es posen en peu. Els ximpanzés poden caminar alguns metres en posició dreta, però ho fan amb molta poca traça. En els hominins bípedes, la major amplada de l’ala ilíaca va canviar el lloc d’inserció dels glutis mitjà i menor, que es van desplaçar des del darrere cap a banda i banda del cos. Amb aquest simple canvi, els dos músculs van modificar la seva funció. L’Homo sapiens s’impulsa gràcies a l’esforç d’adducció del gluti major, però els glutis mitjà i menor són abductors i es tensen cada vegada que aixequem alternativament les cames per caminar, impedint que ens caiguem quan només estem recolzats en una de les dues cames. Els glutis de simis i humans són els mateixos, però la seva funció ha canviat. Quan els nostres avis perden el to muscular dels glutis també necessiten el suport d’un bastó per poder mantenir-se en peu i caminar.
D’altra banda, el nostre os sacre és notablement més ample que en els simis i l’anell pèlvic té una forma diferent. A més, en la nostra espècie els ossos púbics són curts i amples i l’isqui també ha experimentat una reducció significativa (la disposició geomètrica dels tres ossos de la pelvis i l’amplada de l’os sacre té relació amb el part).
A subratllar els músculs isquiotibials (grup muscular situat a la part posterior de la cuixa, estès entre la pelvis i el genoll, format pel múscul semimembranós, el semitendinós i el bíceps femoral), atès que tenen gran importància en el sistema muscular perquè actuen com a extensors dels malucs i flexors del genoll. Però aquestes funcions tenen un límit en la nostra espècie perquè som bípedes, mentre que en els simis no s’esdevé el mateix, ja que són capaços d’una major flexió del maluc, el que els permet enfilar-se amb enorme facilitat. No obstant això, convé recordar que Vivek Venkatamaran[4] i el seu equip han observat els costums enfiladissos de les tribus Twa de les regions equatorials d’Àfrica central. Els membres d’aquestes tribus són capaços de grimpar als arbres flexionant el maluc més de 45 graus, superant amb escreix les habilitats de qualsevol de nosaltres i demostrant la «convivència» entre la bipedestació i la possibilitat de grimpar amb enorme facilitat, encara que no disposem d’elements anatòmics propis de les espècies genuïnament enfiladisses.
La pelvis dels australopitecs, molt més propers en el temps a l’antecessor comú que a nosaltres, té totes les característiques biomecàniques i arquitectòniques necessàries perquè aquells hominins caminessin de manera bípeda. Sembla obvi que els canvis van haver d’esdevenir en un mitjà mancat del perill que suposaven els predadors i en un temps relativament breu.
Els nostres peus també han experimentat un canvi molt notable. Les petjades de Laetoli, a Tanzània, demostren que fa 3,7 milions d’anys la forma del peu dels hominins ja era similar a la d’Homo sapiens. El peu humà ha hagut de modificar-se per suportar tot el pes del cos, deixant enrere la possibilitat d’una gran capacitat per a la flexió, que permet habilitats enfiladisses: es tracta de dues veritables plataformes, que poden suportar un cos situat en posició vertical i ajudar a impulsar el nostre desplaçament; des de fa almenys 3,7 milions tenim un primer dit (hallux) alineat amb els altres dits; també disposem de tres arcs plantars característics: dos d’ells tenen una disposició longitudinal, l’intern és més alt que l’extern, i l’arc transversal està format pels ossos metatarsians. La volta plantar formada per aquests arcs té tres punts de suport. Aquesta volta proporciona estabilitat i resistència, dissipa les fortes càrregues que s’exerceixen sobre el peu i economitza energia quan caminem o correm.
La planta del peu per sota de l’os calcani (taló) està cobert per una gruixuda capa de teixit connectiu subcutani format per fibres de col·lagen, que conté càmeres de pressió. Aquest teixit tan especial actua com a amortidor de l’impacte del peu contra el terra quan suporta tot el pes del cos. En realitat, la tecnologia de les sabatilles dels corredors de marxa no fa sinó imitar la natura amb les seves càmeres d’aire incorporades.
El pes del cos, tant en posició estàtica com durant la marxa, arriba fins al turmell, format per l’astràgal i les epífisis de peroné i de la tíbia, i fins al calcani. Des d’aquests elements ossis la càrrega arriba fins als dits dels peus seguint eixos externs i interns. El conjunt suporta les enormes càrregues que s’exerceixen sobre el peu i actua com a amortidor durant la marxa. A més, el peu té una funció motora, impulsant el cos cap a davant, i una funció d’equilibri fonamentada sobretot en els elements ossis i lligaments del turmell. En definitiva, el peu d’Homo sapiens i el de tots els nostres avantpassats representa un conjunt d’ossos, músculs i lligaments extraordinàriament especialitzat i molt diferent de les potes del darrere dels mamífers.
2. Hipòtesis sobre l’origen del bipedisme
a) La interpretació «clàssica»: té a veure amb els canvis ambientals que es van desenvolupar a l’Àfrica i en altres parts del globus fa entre set i vuit milions d’anys. En aquell moment es va produir un important canvi en l’estructura de la vegetació a nivell global, de manera que els arbres i arbustos (que utilitzen la ruta fotosintètica coneguda com C3) van cedir el seu domini a les prades herbàcies (que utilitzen la ruta fotosintètica coneguda com C4). Aquest canvi va coincidir amb un significatiu refredament global i amb una nova expansió dels gels a l’Antàrtida (i, potser també, per primera vegada, a l’Àrtic). A l’Àfrica, aquest canvi es va traduir en una regressió dels boscos i una primera expansió de les sabanes, amb tota la seva fauna associada. En aquest context, el bipedisme hauria nascut com una resposta a les noves condicions. La locomoció bípeda hauria permès a aquells homínids que es van allunyar del bosc sobreviure en el nou mitjà de la sabana. Entre altres coses, la postura bípeda els hauria permès atalaiar per sobre de les altes herbes i els matolls que són dominants en alguns ecosistemes africans. Així mateix, en un context de progressiva desforestació, l’adopció d’una postura bípeda hauria mitigat els efectes nocius d’una exposició prolongada als raigs del sol, en reduir la superfície corporal exposada verticalment a aquest. Igualment, en el context de migracions estacionals que és comú a bona part de la fauna de la sabana actual, podem imaginar que una locomoció bípeda obligada i no ocasional (com en el cas dels bonobos), hauria afavorit així mateix la supervivència d’aquests homínids en un entorn canviant, obligats a trobar nous hàbitats favorables i a desplaçar-se periòdicament. La locomoció bípeda hauria aparegut com una resposta eficaç a aquest règim climàtic estacional.
Aquesta hipòtesi d’un primat que es va aixecar sobre les seves cames per guanyar en alçada, visualitzant així les preses en les sabanes per sobre de l’alçada de les diferents espècies de gramínies (que poden assolir o superar els dos metres), actualment es considera una simple anècdota.
b) Ha estat sobretot Owen Lovejoy, membre de l’equip que va descriure a Lucy i els primers Australopithecus afarensis, el que ha aportat els arguments més incisius a l’hora de deslligar el bipedisme del desenvolupament de les prades herbàcies. Per Lovejoy, el bipedisme, i el consegüent alliberament de les mans, es va seleccionar pel seu paper en l’estratègia reproductora de l’espècie. Segons aquest autor, el nou tipus de locomoció hauria d’haver permès als mascles el transport de recursos suplementaris amb els quals mantenir a les femelles i les seves cries, la qual cosa al seu torn permetria a aquestes procrear més assíduament i augmentar l’eficàcia reproductora d’aquests primers homínids . El bipedisme també hauria permès a les femelles carregar amb les seves cries mentre realitzaven altres tasques en cooperació amb els mascles, com és la recol·lecció de llavors.
Existeixen, a més, altres arguments que reforcen els punts de vista de Lovejoy, com ara la capacitat per escapar amb les cries i així salvaguardar-se dels depredadors. És sabut que per a qualsevol mamífer i, per descomptat, per a qualsevol primat, els elements més exposats a la depredació són precisament les cries. En els primats quadrúpedes i també en els nostres germans antropomorfs, poc després de néixer les cries poden valer-se per si mateixes i, per tant, escapar d’un possible depredador sense dependre de la mare. Però l’alliberament de les mans va proporcionar un nou mecanisme de seguretat per a les cries. Aquestes podien escapar dels depredadors en braços de les seves mares o pares. Això comportaria una major dependència dels progenitors i un estrenyiment de la relació mare-fill, afavorint el contacte continuat i els mecanismes d’aprenentatge. Potser pogués també comportar un estrenyiment de les relacions entre els germans, el que comportaria l’estructuració de grups familiars.
Aquesta hipòtesi, proposada en els anys 1960 i 1970 per Owen Lovejoy o Robert Ardrey, que incidia en els avantatges que hauria suposat disposar de les mans per fabricar i transportar objectes, va tenir molta acceptació i va resistir una mica més d’una dècada. S’assumia que la bipedestació hauria sorgit com a resposta a la necessitat d’alliberar les mans. La pressió selectiva capaç de generalitzar aquesta forma de locomoció estaria, doncs, en la tecnologia i, com una conseqüència addicional favorable, la possibilitat de transportar les cries o portar menjar a altres membres del grup hauria reforçat la bipedestació.
Però les troballes realitzades per Donald Johanson i Timothy White al jaciment de Hadar (Etiòpia) durant la dècada de 1970 van permetre rebutjar aquesta hipòtesi. L’anatomia de l’Australopithecus afarensis i les petjades de Laetoli, d’una antiguitat d’un 3,8 milions d’anys, van certificar que fa més de 3 milions d’anys aquests hominins havien caminat drets i, en aquells moment, no existien dades fefaents per argumentar que aquesta espècie havia fabricat eines.
c) descartada la hipòtesi de l’ús de les mans «alliberades» de la locomoció, i sabent que els australopitecs de l’est d’Àfrica van viure en ambients de sabana, Peter Wheeler va pensar llavors en els possibles avantatges o desavantatges d’un primat bípede sotmès als rigors climàtics d’ambients mancats de la protecció de la cobertura vegetal. Després d’un exhaustiu estudi sobre la fisiologia de la termoregulació, Wheeler va concloure que un primat bípede pot ser més eficient que un quadrúpede des del punt de vista de la regulació de la seva temperatura corporal en els ambients oberts tropicals i subtropicals. Arran de terra l’aire és més càlid i està en calma. Els quadrúpedes exposen més superfície corporal a la intempèrie i perden per això més quantitat d’aigua. En conseqüència, un primat bípede necessita menys aliments i menys aigua per desplaçar-se, encara que ho faci amb menys rapidesa que un quadrúpede a la cursa. Duncan Mitchell i el seu equip de treball van demostrar que els babuins resisteixen molt bé la vida en les sabanes, excepte quan l’aigua escasseja, i van recolzar la hipòtesi de Wheeler amb la idea que les mans lliures dels hominins podrien haver estat utilitzades per transportar aigua en moments d’escassetat. Però George Chaplin i el seu equip també han realitzat experiments, en els quals demostren una eficàcia energètica similar en els ambients oberts i càlids de les sabanes africanes, tant si se és bípede com quadrúpede. També Karen Steudel-Numbers, estudiant els costos energètics que comporta la locomoció, va concloure que la bipedestació no és més eficient que la locomoció quadrúpeda. I Michael Sockol i els seus col·laboradors van arribar a la conclusió que la marxa bípeda requereix un 75% menys d’energia que la marxa quadrúpeda, un descens atribuïble a la forma del nostre maluc i a la longitud dels membres inferiors.
d) per a altres autors l’explicació que lliga l’origen del bipedisme humà a l’adaptació de la vida a la sabana no és exacta. Existeixen arguments per pensar que l’adopció del bipedisme es va produir en un medi boscós i no a la sabana. Així, l’anàlisi detallada de l’anatomia de Lucy per part de Randall L. Susman indica que aquest homínid, malgrat el seu bipedisme, mostra trets que el vinculen encara a una vida associada al bosc. Per exemple, les falanges de mans i peus d’Australopithecus afarensis eren corbades, com passa en tots els primats que acostumen a agafar-se a les branques. D’altra banda, les primeres anàlisis d’un esquelet molt complet d’Australopithecus trobat per Ron Clarke el 1997 a la localitat sud-africana de Sterkfontein indiquen així mateix una vida més associada al bosc que a la sabana. Encara que l’extracció de l’esquelet de Sterkfontein no ha estat completada, els petits ossos del peu d’aquest esquelet han proporcionat una informació sorprenent. En articular el primer metatars amb el corresponent os del tars (l’anomenat primer cuneïforme), Ron Clarke i Philip Tobias van arribar a la conclusió que aquest havia de formar amb la resta dels dits un angle més obert del que s’esperava per a un homínid típicament bípede . En altres paraules, l’Australopithecus del Membre 2 de Sterkfontein presentava un dit gros (o hàl·lux) encara abduït i separat de la resta dels dits del peu, retenint una funció prensora semblant a la que s’observa en ximpanzés i goril·les. Per Clarke i Tobias, això provava que els primers Australopithecus haurien portat una vida fonamentalment arborícola, ajudant-se de les seves mans i peus per pujar als arbres, tal com passa avui dia en els antropomorfs vivents.
Com era el medi en el que varen viure els més remots avantpassats?
Quan Wheeler va publicar els seus treballs, allà per la dècada de 1990, a l’Àfrica es produïen noves troballes d’hominins encara més antics que els propis australopitecs. Timothy White i un extens equip de col·laboradors van començar a obtenir fòssils d’una nova espècie en jaciments d’Etiòpia. El 1994 es van conèixer les primeres dades sobre Ardipithecus ramidus, que va viure fa entre 4,5 i 4 milions d’anys. Aquesta espècie podia ser descendent d’una altra encara més antiga, Ardipithecus kadabba, descoberta per Yohannes Haile-Selassie també a Etiòpia, i amb una cronologia que s’acosta als 6 milions d’anys: és a dir, gairebé en el límit proposat pels genetistes per a la separació de les respectives genealogies de ximpanzés i humans. No sembla haver-hi dubtes sobre el bipedisme d’Ardipithecus ramidus. En cas de demostrar-se de manera definitiva que les dues espècies van ser bípedes i que no van habitar en mitjans oberts, la hipòtesi de Wheeler quedaria rebutjada. Què en sabem, doncs, de l’hàbitat dels ardipitecs?
Aquests fòssils estan associats a més de 60 espècies de vertebrats, que majoritàriament suggereixen un ambient humit i boscós, característics de riberes de riu i llacs, faltant altres típiques de prades. No obstant això, la presència de bòvids també suggereix l’existència de pastures en ambients més oberts. En resum, tot apunta que aquesta espècie bípeda va viure en ambients de boscos tancats, en els quals les branques més altes s’entrellaçaven i formaven un ambient humit i gairebé desproveït de llum solar directa, sense descartar la possibilitat que aquells hominins s’endinsessin en camp obert.
El medi on va viure l’espècie Ardipithecus ramidus, un milió d’anys més recent que Ardipithecus kadabba, suggereix un medi humit de bosc, boscos tancats, però amb una dieta molt diversa i pròpia d’un primat omnívor, atès que no té les especialitzacions dels simis, que tendeixen a una dieta frugívora i folívora.
Però aquestes conclusions han estat replicades per Thure Cerling i un ampli grup d’experts; emprant la mateixa informació aportada pels investigadors de l’equip de White, l’equip de Cerling considera que l’hàbitat d’Ardipithecus ramidus consistia més aviat en una regió coberta per un 25% de zones boscoses, com a màxim, corresponent la resta a zones obertes de sabana amb herbàcies i matolls.
L’anatomia del peu d’Ardipithecus ramidus, que es coneix molt bé, no pot mostrar l’ambient en què van viure aquests hominins, però permet realitzar algunes inferències molt interessants. D’una banda, un seguit d’elements anatòmics similars a la d’altres primats arborícoles, ens parla d’un medi forestal. D’una altra, l’anatomia dels metatarsians d’Ardipithecus suggereixen immobilitat plantar en lloc de la flexibilitat del peu dels ximpanzés o dels goril·les, cosa que fa pensar que el peu d’Ardipithecus ramidus es mantenia ferm quan els individus caminaven o corrien. I pel que fa a la seva pelvis, l’escurçament, amplada i geometria de l’ili indiquen clarament la disposició dels glutis per impulsar el cos cap endavant (gluti major) i per l’abducció del maluc durant la bipedestació (glutis mitjà i menor). En canvi, la morfologia de l’isqui i del pubis encara presenta similituds amb la dels simis, que permetrien una notable acció dels músculs isquiotibials en la flexió del maluc. Ardipithecus ramidus tindria, per tant, una gran facilitat per grimpar. En definitiva, aquesta espècie presenta un mosaic anatòmic perfecte per desplaçar-se sobre terra amb les cames i per moure’s per les branques en un ambient de bosc. És molt probable que Ardipithecus ramidus no pogués córrer com ho fem nosaltres. Si és així, és molt possible que els membres d’aquesta espècie no s’arrisquessin a tenir aventures duradores en espais oberts ja que no podien competir amb els predadors. És per això que la balança sembla inclinar-se cap la hipòtesi de l’inici de la bipedestació en ambients de boscos tancats.
Conclusions
L’escassa informació disponible ens condueix a les següents conclusions:
- és molt possible que el nostre avantpassat comú no tingués una locomoció similar a la de goril·les i ximpanzés. Aquests simis haurien evolucionat cap a la seva particular manera de desplaçament actual, recolzant els artells a terra.
- sembla probable que el nostre avantpassat comú amb els ximpanzés fos bípede.
- la bipedestació va poder sorgir en un ambient de bosc tancat i no en camp obert, com s’ha suggerit de manera repetida en dècades anteriors.
- el nostre avantpassat comú va poder tenir una altura i un pes probablement similars als dels ximpanzés. Aquesta dada ens porta a pensar que part de la seva vida es va desenvolupar a terra dels boscos africans, sense descartar que fos capaç de grimpar amb gran facilitat a les branques més resistents a la recerca de menjar o refugi.
- si tot el que portem dit és correcte, es podria acceptar que el nostre avantpassat comú amb els ximpanzés va trobar aliment abundant tant a terra dels boscos com durant les seves aventures de curta durada i esporàdiques en camp obert. D’aquesta manera, aquest ancestre no va entrar en competència amb altres primats arborícoles, en diversificar el seu aliment cap a una dieta més omnívora.
La forma de caminar bípeda, combinada amb la facilitat per grimpar, va poder sorgir en ambients tancats sense pressions selectives considerables, com una forma més de desplaçament, que es va fer cada vegada més necessària a mesura que es produïa la regressió de la coberta vegetal. Els avantpassats dels ximpanzés van ocupar el nínxol ecològic d’un primat de la seva envergadura, molt similar al de l’ancestre comú, mentre que els avantpassats del nostre llinatge van tenir la possibilitat de sortir cap a camp obert i ocupar un nínxol diferent. Només així podem explicar la divergència genètica de les dues genealogies, que no trigarien a ocupar regions molt allunyades. Molt més tard, la bipedestació va oferir avantatges termoreguladors i la capacitat per utilitzar les mans en funcions diferents, com la fabricació d’eines. És el que en biologia evolutiva es coneix com exaptació, un terme introduït en 1982 per Stephen Jay Gould, que es refereix a caràcters que ofereixen una capacitat adaptativa diferent de l’original, molt de temps després de la seva aparició en una espècie.
En aquestes primeres etapes de locomoció bípeda, sembla que tenim uns homínids anatòmicament partits en dos de manera que l’evolució del maluc cap amunt es dissocia de l’evolució del maluc cap avall. Maluc cap amunt trobem caràcters com és la longitud relativa dels braços o la forma de les falanges, que semblen associats encara a un hàbitat boscós. Per contra del maluc cap avall, les cames tendeixen a allargar-se i a adoptar una marxa bípeda cada vegada més eficaç.
Lectures de referència
Agustí, Jordi; Bufill, Enric i Mosquera, Marina (2012). El precio de la inteligencia. Barcelona. Crítica. (Col. Drakontos). 256 pàgs.
Allman, John Morgan (2003). El cerebro en evolución. Barcelona. Ariel. (Col. Ariel neurociencia). 1ª impr. 240 pàgs.
Arsuaga, Juan Luis i Martínez, Ignacio (2006). La especie elegida : la larga marcha de la evolución humana. Madrid. Ed. Temas de Hoy. 1ª impr. 448 pàgs.
Briones, Carlos; Fernández, Alberto i Bermúdez, José María (2015). Orígenes. El universo, la vida, los humanos. Barcelona. Crítica. Pàgs. 341-469
Campàs, Joan (2014): 15. Els homínids: els preaustralopitecins. Curs en línia: «Els orígens de l’art i l’evolució humana: l’Homo significans». En línia a http://cv.uoc.edu/adf/~04_999_01_u07/llico_15.pdf
Carbonell i Roura, Eudal (2005). Homínidos : las primeras ocupaciones de los continentes. Barcelona. Ariel. (Col. Ariel prehistoria ). 864 pàgs.
Leakey, Richard Erskine (2000). El origen de la humanidad. Barcelona. Debate. (Col. Temas de Debate). 1ª impr. 224 pàgs.
Striedter, Georg F. (2005). Principles of brain evolution. Sunderland MA. Sinauer Associates. 436 pàgs.
Notes
[1] Morris, Desmond (2006). El mono desnudo. Barcelona. Debolsillo ed (Random House Mondadori). (Col. Ensayo, 58). 5ª Impr. 272 pàgs.
[2] Grups taxonòmics: Domini, Regne, Subregne, Branca, Infraregne, Superfílum / Superdivisió, Fílum / Divisió, Subfílum / Subdivisió, Infrafílum, Microfílum, Nanofílum, Superclasse, Classe, Subclasse, Infraclasse, Parvclasse, Superlegió, Legió, Sublegió, Infralegió, Supercohort, Cohort, Subcohort, Infracohort, Magnordre, Superordre, Grandordre, Mirordre, Ordre, Subordre, Infraordre, Parvordre, Superfamília, Grup, Família, Subfamília, Infrafamília, Supertribu, Tribu, Subtribu, Infratribu, Gènere, Secció, Subsecció, Sèrie, Espècie, Subespècie, Varietat, Soca
[3] Michel Brunet i altres: «Geology and palaeontology of the Upper Miocene Toros-Menalla hominid locality, Chad». Nature 418, 152-155 (11 July 2002). En línia a http://www.nature.com/nature/journal/v418/n6894/full/nature00880.html
[4] Vivek V. Venkataraman, Thomas S. Kraft i Nathaniel J. Dominy: «Tree climbing and human evolution». PNAS, 2912, vol. 110 no. 4 , 1237–1242 En línia a http://www.pnas.org/content/110/4/1237.full
Dr. Joan Campàs Aura digital
Estudis d’Arts i Humanitats de la UOC
Curs Som les nostres connexions
Material en format pdf
Tema 0. Presentació i justificació
Tema 1. El cervell ens enganya
Tema 2. El cervell també enganya l’autopercepció del cos
Tema 3. Anatomia cerebral bàsica
Tema 4. Mite: Només utilitzem el 10 % del nostre cervell
Tema 5. Mite: L’homosexualitat (l’hetero, la bi, la trans) és una elecció
Tema 6. Mite: Els fills hereten la intel·ligència dels pares
Tema 7. Mite: Internet i els videojocs són addictius
Tema 8. Mite: El cervell és un ordinador
Tema 9. La memòria
Tema 10. Com definir el «jo»? El cervell i la «joïtat»
Tema 11. L’evolució del cervell (1)
Tema 12. L’evolució del cervell (2)
Tema 13. L’evolució del cervell (3)
Tema 14. L’evolució del cervell (4)
Tema 15. Cervell humà i evolució
Conjunt dels posts amb les introduccions fetes al facebook
Som les nostres connexions