Cervell humà i evolució (som les nostres connexions 15)
9 agost, 2016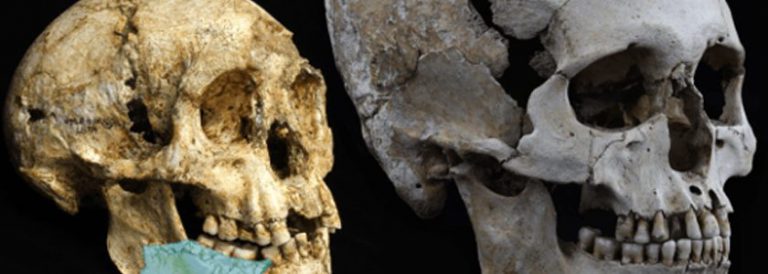
L’home és home pel seu cervell i va obtenir el seu cervell
per ser un primat i descendent de primats.
William W. Howells
Reflexió inicial
Homo sapiens és l’única espècie de la genealogia humana que queda al planeta. Tots els nostres ancestres han desaparegut i aquest fet ens fa reflexionar sobre moltes qüestions. És obvi preguntar-se per la nostra possible superioritat cognitiva sobre les altres espècies, que van anar desapareixent a mesura que ens propagàvem pels cinc continents. Un exercici interessant podria consistir a comparar les eines i el comportament (inferit del registre arqueològic) dels neandertals i dels sapiens arcaics de cronologies similars. Paola Villa i Wil Roebroeks [1] arriben a la conclusió que no hi ha diferències apreciables en l’utillatge de les dues espècies que justifiquin la superioritat tecnològica de l’Homo sapiensdavant Homo neanderthalensis. Cap de les hipòtesis sobre la inferioritat en la capacitat innovadora, cinegètica o tecnològica dels neandertals pot resistir les proves de contrast a partir de les evidències del registre arqueològic. Els neandertals es comunicaven mitjançant un llenguatge propi, tenien capacitat simbòlica i una enorme resistència als rigors climàtics. Per a aquests investigadors potser caldria tornar a replantejar l’assimilació genètica dels neandertals, ara que els genetistes han provat la possibilitat de mestissatge amb descendència fèrtil entre els dos tipus humans. De fet, tots els representants de la nostra espècie hem mantingut alguns gens de neandertals.
És cert que les poblacions humanes recents hem arribat a nivells culturals que superen amb escreix als de qualsevol altra espècie del passat. No obstant això, aquest fet ha succeït una vegada que van desaparèixer la majoria dels nostres competidors. Potser la forma (que no la mida) del cervell pugui donar-nos alguna resposta. Tot i que alguns anatomistes, com Emiliano Bruner, no han trobat raons per pensar que la forma del cervell tingui cap influència en les capacitats cognitives de les últimes espècies que han habitat el planeta.
La història evolutiva d’Homo sapiens és encara molt breu. No obstant això, en pocs mil·lennis hem aconseguit cotes increïbles de desenvolupament. No donem crèdit als avenços tecnològics produïts, que som incapaços d’assimilar en el dia a dia. Què està passant amb la nostra espècie?, què tenim de peculiar?, per què estem aconseguint desenvolupaments cientificotècnics tan impressionants en uns pocs decennis? ¿Potser aquest èxit evolutiu ha tingut a veure amb la complexitat del llenguatge? Certament, el llenguatge és una eina biològica i cultural molt poderosa, però no pot ser l’única. Si ens fixem en el cervell dels humans, podem pensar en les subtils diferències de forma en relació als altres primats. Aquestes diferències no semblen massa significatives ni concloents. La mida del cervell tampoc sembla una resposta adequada. L’única distinció anatòmica amb els nostres avantpassats de finals del Plistocè Mitjà és la grandària relativa del cerebel, que possiblement va augmentar el seu volum en els últims 60.000 anys. A grans trets, podem descartar diferències substancials entre el cervell dels primers membres d’Homo sapiens i nosaltres mateixos. D’altra banda, l’única via que tenim per accedir a la ment dels primers sapiens és el registre arqueològic, que ens parla de capacitats cognitives i de comportament. Però les dades són insuficients. En qualsevol cas, fa 150.000 anys érem caçadors i recol·lectors, la domesticació dels animals i l’agricultura han arribat fa uns pocs mil·lennis, la revolució industrial data de finals del segle XVIII, l’exploració espacial és una fita del segle XX. El model encaixa amb un progrés exponencial de la tecnologia i suscita moltes preguntes.
Quines van ser les causes que van promoure la transformació d’un cervell no molt diferents a les d’un simi en un cervell humà capaç de llenguatge i raonament? Quins canvis genètics, funcionals i estructurals van produir el cervell humà?
Sobre l’origen d’Homo sapiens: la hipòtesi multiregional i la de l’origen africà
L’origen de la nostra espècie és un tema sobre el qual ja va reflexionar Charles R. Darwin, després de posar les bases de l’evolució de les espècies per selecció natural. Un segle després, l’aparició dels humans moderns ha estat potser el debat més apassionant dels últims 30 anys. No és per a menys, ja que en paral·lel a les dades empíriques obtingudes sobre aquesta qüestió s’ha desenvolupat una polèmica que transcendeix la ciència i interessa a filòsofs, religiosos i defensors d’idees més o menys passades, quan no perilloses. Estem preguntant-nos per nosaltres mateixos. Volem saber com, quan i per què va sorgir Homo sapiens. No són qüestions banals, que podrien ser enfocades des de molts punts de vista.
L’anatomista alemany Franz Weidenreich (1873-1948) va tenir oportunitat d’estudiar tot el registre fòssil de la seva època, que es limitava fonamentalment a les troballes produïdes a Euràsia. Gràcies a les seves investigacions amb els fòssils trobats a la Xina, i en particular els del jaciment de Zhoukoudian, Weidenreich va ser el precursor de la teoria multiregional sobre l’origen de les poblacions actuals. Segons ella, tots els éssers humans procediríem de l’evolució d’una espècie, que va habitar al Vell Món i les poblacions mai van deixar de perdre el contacte genètic entre elles. El resultat va ser el sorgiment d’Homo sapiens, una mena de gran diversitat morfològica. Sense entrar en detalls i en el debat corresponent, és evident que la cultura de finals del segle XIX i la meitat del segle XX estava encara impregnada d’un fort sentiment racista. Els éssers humans del planeta no tenim la mateixa aparença i les diferències s’accentuen mitjançant elements culturals, com l’abillament, els objectes simbòlics que adornen el cos i els propis costums.
Carletoon S. Coon (1904-1981) va ser un antropòleg americà, fortament influït per la teoria poligènica sobre l’origen de la humanitat en races diferents, així com pels treballs de Weidenreich. Per a Coon, l’existència de races és un fet provocat per l’origen geogràfic de cadascuna d’elles. La rivalitat entre totes les poblacions humanes del planeta hauria portat a una selecció de la diversitat actual. Les observacions de Coon prediuen l’existència de fins a cinc races diferents i la ferma convicció de la superioritat d’unes sobre les altres. La seva obra dedicada a les races d’Europa, publicada el 1939, va ser el preludi de la seva teoria sobre la superioritat de certs grups humans. Dissortadament, la influència de l’obra de Coon ha arribat fins als nostres dies: les teories racistes segueixen vives, tot i l’enorme quantitat de dades empíriques en contra. De fet, la seqüenciació del genoma humà ha demostrat que, en realitat, els gens ens diuen que les races no existeixen.
El 1984 els paleoantropòlegs Milford H. Wolpoff (Universitat de Michigan, USA), Alan S. Thorne (Universitat Nacional d’Austràlia) i Wu Zinzhi (Institut de Vertebrats i Paleontologia de Pequín) van iniciar una defensa a ultrança de la teoria multiregional de la nostra espècie postulada per Franz Weidenreich. Fins a la dècada de 1980, l’autoritat moral que Ernst Mayr (1904-2005) va imposar el 1950 sobre la taxonomia dels hominins només va tenir resposta en la gosadia de Richard Leakey (1944), Philip Tobias (1925-2012) i John Napier, quan en 1964 van proposar l’espècie Homo habilis. Però l’entrada en escena d’aquesta nova espècie no va entelar la idea que Homo erectus va ser la gran conqueridora del Vell Món. Wolpoff, Thorne i Wu han seguit la taxonomia sintètica de Mayr i han defensat sempre la unicitat d’Homo erectus, l’antiguitat del qual es xifra ja en almenys 1,8 milions d’anys. Aquesta espècie s’hauria expandit per Àfrica, Àsia i Europa, però les seves poblacions haurien mantingut un contacte genètic a través del temps. Com a resultat de l’evolució d’Homo erectus, la nostra espècie va anar apareixent a l’Àfrica i Euràsia a través de formes intermèdies, cada vegada més semblants a nosaltres. Wolpoff, Thorne i Wu no han defensat l’evolució paral·lela de poblacions distanciades per milers de quilòmetres, sinó l’evolució d’una espècie cosmopolita estretament unida per llaços genètics. Els exemplars d’aspecte més evolucionat, com les restes cranials de Dali, Jinniushan i Maba (Xina) representarien formes intermèdies entre Homo erectus i Homo sapiens i la millor prova que la hipòtesi de l’origen multiregional de la nostra espècie és encertada. La diversitat que avui podem observar entre diferents poblacions seria el resultat d’adaptacions locals i no d’un origen poligènic de fortes connotacions racials.
No obstant això, l’increment del registre fòssil ha estat inexorable i el progrés en l’àmbit de la genètica ha ofert noves respostes. Abans d’exposar la hipòtesi alternativa a l’origen multiregional cal recordar que la densitat demogràfica de les espècies del Plistocè va ser molt baixa i el model de la desaparició continuada de grups humans va poder ser la norma més que l’excepció. Àfrica i Europa van tenir barreres geogràfiques inexpugnables motivades per la successió d’èpoques glacials i interglacials, d’intensitat creixent durant l’últim milió d’anys. Les investigacions en l’àmbit de la genètica corroboren el distanciament evolutiu entre poblacions que van viure separades durant milers d’anys. Els neandertals són l’exemple millor conegut. Cada vegada resulta més complicat defensar la integritat d’una espècie humana en el Plistocè, distribuïda per una extensió de prop de 85 milions de quilòmetres quadrats, compartimentada per la Mediterrània, dotzenes de cadenes muntanyoses i deserts inhabitables. Els suposats llaços genètics semblen més una quimera que una realitat en el món perdut del Plistocè.
Així i tot, molts científics segueixen defensant que l’espècie Homo erectus va tenir una gran longevitat i que va ocupar una gran extensió territorial d’Àfrica i Euràsia. Però, amb independència de debats i confrontacions sobre el nom que hagi de rebre cada població del Plistocè, són ja molt pocs els que pensen en una transformació evolutiva d’Homo erectus cap a Homo sapiens. La teoria multiregional s’ha anat diluint en el mar de dubtes generat per les evidències del registre fòssil i per l’avanç de les investigacions sobre el genoma humà i el d’algunes poblacions pretèrites.
A mesura que s’han anat produint descobriments a l’Àfrica i Euràsia, les evidències ens han mostrat un panorama evolutiu molt suggeridor durant el Plistocè Mitjà. Mentre que a Europa es desenvolupaven els avantpassats dels neandertals, d’aspecte inconfusible, en la resta d’Euràsia evolucionaven poblacions amb un patró molt general de trets arcaics. Àfrica, en canvi, sembla ser l’escenari d’una història molt diferent. Els fòssils africans de la segona meitat del Plistocè Mitjà són molt particulars. Podem destacar els d’Eliye Springs [2] i KNM-ER 3884 (Etiòpia), Florisbad i KIasies Ríver Mouth (Sud-àfrica), Jebel Irhoud (Marroc), Herto i Omo-Kibish (Etiòpia) i Ngaloba 18 (Tanzània). Aquests fòssils semblen estar explicant una història diferent a la d’Euràsia. La seva antiguitat és inferior a 300.000 anys i el seu aspecte, de totes totes modern, està clarament derivat cap a la morfologia de la nostra espècie. En particular, les restes cranials del jaciment de Herto han estat assignats a la subespècie Homo sapiens idaltu per Timothy White, pel fet que no és possible distingir-los d’espècimens robustos de la nostra espècie. L’exemplar BOU-VP-16/1 està representat per un crani alt i arrodonit, amb un volum intern de 1.450 centímetres cúbics així com per una cara robusta però idèntica a la nostra. Les datacions mitjançant el mètode dels isòtops de l’argó per les restes de Herto estimen l’antiguitat d’aquesta població en uns 160.000 anys. No hi ha dubte que al continent africà s’estava produint un procés evolutiu diferent al que succeïa a Euràsia. Com explicar l’aspecte gairebé plenament «sapiens» de les poblacions africanes de la segona meitat del Plistocè Mitjà pel que fa a les eurasiátiques? Sota la perspectiva de la hipòtesi multiregional, els fòssils africans d’aquesta època representen un greu problema.
L’esquema presenta les dues hipòtesis que han competit durant anys sobre l’origen d’Homo sapiens. El model multiregional proposa una continuïtat evolutiva a Àfrica, Àsia i Europa des de la primera expansió dels hominins fora d’Àfrica. En l’actualitat s’assumeix que aquesta expansió va succeir fa uns dos milions d’anys. Les poblacions que van viure tant a l’Àfrica com a Euràsia haurien mantingut un cert contacte, evitant així processos d’especiació. D’aquesta manera i de manera progressiva, l’espècie Homo erectus hauria donat lloc a la nostra espècie fa uns 100.000 anys. El model d’origen africà postula l’aparició d’Homo sapiens a l’Àfrica fa uns 200.000 anys. Després de la seva expansió fora del continent africà, la nostra espècie va substituir a les poblacions d’Homo erectus, Homo neanderthalensis, etc., que habitaven a l’Àfrica i Euràsia. Les investigacions sobre el genoma de poblacions recents i en alguns fòssils donen suport a aquest últim model, sense oblidar que aquestes investigacions reconeixen ara la possibilitat d’hibridacions puntuals d’Homo sapiens amb les poblacions autòctones de cada continent.
Els paleoantropòlegs Günter Brauer i Christopher Stringer van ser els primers a oferir una solució a aquest aparent problema. L’espècie Homo sapiens es va originar a l’Àfrica i només en aquest continent, potser mitjançant un coll d’ampolla en una població de pocs membres efectius. Euràsia hauria quedat llavors al marge del procés evolutiu que va donar lloc a la nostra espècie. Aquesta hipòtesi va sorgir a meitat de la dècada de 1980: la teoria multiregional havia estat qüestionada per primera vegada. El canvi de paradigma estava en potència. En poc temps la hipòtesi alternativa va ser coneguda com Out Of Àfrica.
El problema sobre l’origen de la nostra espècie era massa important com per quedar-se només en l’àmbit de la paleoantropologia. Els genetistes van entrar amb força i enginy en la qüestió. El mtDNA africà representaria la seqüència primitiva de la qual haurien derivat les altres. El temps estimat perquè el mtDNA ancestral s’hagués diversificat en les variants actuals es va valorar (considerant una taxa de mutació estàndard) en uns 200.000 anys. Ja que les mitocòndries i, per tant, el mtDNA procedeixen de l’òvul i no dels espermatozoides, la conclusió d’un article pioner [3] suggeria que les primeres dones de la nostra espècie van aparèixer a l’Àfrica durant el Plistocè Mitjà, coincidint amb les dades aportades pel registre fòssil.
Com bé podem imaginar, els resultats de Cann i els seus col·legues van representar un revulsiu molt important en les investigacions sobre el nostre origen. Molt aviat aquells resultats van ser batejats com la teoria de la «Eva mitocondrial» o la «Eva Negra», en al·lusió al fet que les dades es referien a dones i no a homes. I aquest va ser precisament un dels principals esculls que van trobar Cann, Stoneking i Wilson. Es pot explicar un fet tan crucial per a la humanitat estudiant tan sols el mtDNA?, què va succeir amb els homes? Aquesta última qüestió es va haver de resoldre com més aviat millor, perquè la història evolutiva de la nostra espècie ha de ser la mateixa per als dos sexes. La resposta va poder trobar-se estudiant el DNA nuclear, que compartim tots els humans (i que evoluciona més lentament que el mitocondrial). Però, en particular, els experts es van fixar en el cromosoma Y, i les investigacions del DNA nuclear del cromosoma Y van portar a les mateixes conclusions a què havien arribat Cann i els seus col·legues el 1987 amb el mtDNA. L’origen de la nostra espècie es localitza a l’Àfrica sub sahariana, des d’on es va dur a terme una gradual expansió cap als confins del planeta durant els últims 100.000 anys. La hipòtesi de l’Out Of Africa implica una única expansió, ocorreguda fa menys de 100.000 anys, durant la qual s’hauria produït una disminució de la variabilitat genètica i fenotípica. Ara sabem que l’expansió d’Homo sapiens va succeir en diverses etapes i per llocs diferents. No obstant això, la diferència temporal entre cada expansió és tan petita en termes geològics que, a tots els efectes, podem parlar d’un únic esdeveniment de sortida des del continent que ens va veure néixer com a espècie.
El nou paradigma ha estat admès per la majoria dels especialistes, encara que no tots han acceptat la simplicitat de la proposta. Però queden moltes qüestions en l’aire. En primer lloc, podem preguntar pel lloc utilitzat per a l’expansió de la població africana cap a la resta del planeta. Es donava per fet que els humans d’anatomia moderna, descendents de l’Eva Negra, es van trobar amb els neandertals al Corredor Llevantí fa una mica menys de 100.000 anys. En cert moment, les poblacions de la nostra espècie haurien trencat la resistència dels neandertals, per penetrar primer cap a Euràsia i més tard cap a Austràlia i les Amèriques. Hi ha una certesa molt fundada sobre aquest punt, a tenor de registre fòssil d’aquesta regió, amb restes com els dels jaciments de Jebel Qafzeh i Es Skhül. Aquestes troballes i el mateix plantejament de la hipòtesi de l’Out of Africa impliquen una expansió cap al nord de les poblacions sub saharianes de les que suposadament procedim tots els humans. Aquesta expansió implica el pas franc per una barrera suposadament infranquejable: el gran desert del Sàhara.
Les investigacions sobre la geologia de la regió han demostrat la viabilitat de la vida a les regions del Sàhara en diferents moments del Plistocè, amb una elevada pluviositat, formació de grans llacs i cursos fluvials, i una vegetació suficient com per permetre la vida de nombroses espècies de vertebrats. Per exemple, un escenari com aquest es va produir fa aproximadament 135.000 anys. Un altre similar va passar fa uns 85.000 anys. El pas des de l’Àfrica sub sahariana cap al Corredor Llevantí va quedar expedit en aquests i altres moments del Plistocè i l’Holocè.
També s’ha proposat el pas de les poblacions africanes cap a Euràsia a través de l’estret de Bab al-Mandeb. El braç de mar entre la regió actualment ocupada per l’estat de Djibouti i la península d’Aràbia té una longitud d’uns 30 quilòmetres i una profunditat de poc més de 300 metres. Sens dubte, els nostres avantpassats africans van poder creuar aquest estret, a jutjar per l’antiguitat del jaciment de Jebel Faya, localitzat en l’actual estat dels Emirats Àrabs Units i proper a l’estret d’Ormuz. Les eines de pedra trobades en aquest jaciment tenen una antiguitat aproximada de 125.000 anys i són molt similars a les trobades a l’Àfrica en jaciments contemporanis [4]. Els descensos del nivell del mar durant èpoques glacials van poder facilitar el pas pels estrets de Bab al-Mandeb i Ormuz per colonitzar altres regions del sud d’Àsia. Així doncs, es pot plantejar que l’èxode de la nostra espècie va poder succeir en dues o més onades migratòries a través de llocs diferents i en moments diferents. Existeixen dades recents obtingudes per un equip de genetistes liderats per Gisela Kopp en els papions de l’espècie papió sagrat, que viuen a l’est d’Àfrica i a la costa oriental de la península d’Aràbia. Com aquests primats van ser domesticats pels egipcis es va arribar a pensar que les poblacions de papions de la península d’Aràbia van ser transportades per grups humans en dates històriques. No obstant això, les anàlisis del DNA de diferents poblacions de babuïns d’aquestes regions han demostrat que els exemplars de papió sagrat de la península d’Aràbia es van separar dels seus congèneres africans fa almenys 130.000 anys. La xifra coincideix bé amb el possible pas de la nostra espècie per Bab el-Mandeb i ens convida a pensar que tant els papions com nosaltres vam tenir un pas franc per la banya d’Àfrica abans que acabés el Plistocè Mitjà. Potser els neandertals ens van tancar el camí cap a Europa, on no vàrem arribar fins fa poc més de 40.000 anys. En canvi, els jaciments de la Xina suggereixen la presència d’Homo sapiens més enllà de la barrera dels 100.000 anys (jaciment de Zhirendong, sud de la Xina), un esdeveniment que encara requereix investigació i dades més consistents.
Els cranis africans d’aspecte modern mostren una gran variabilitat i tenen un notable semblança amb poblacions modernes de procedències diverses. Aquest fet suggereix que les poblacions africanes de finals del Plistocè Mitjà no formaven un grup uniforme i que la seva herència està dispersa per tot el planeta. Es poden, doncs, proposar diverses migracions des d’Àfrica, en un període que va començar al voltant de fa 130.000 anys, com recolzen les dades del jaciment de Jebel Faya. La colonització del planeta per Homo sapiens es va produir des d’Àfrica, però el model pot ser molt més complex del que suggereixen les primeres dades ofertes per la investigació del genoma humà.
Augment de mida del cervell
Increment de la mida del cervell en els últims 7 milions d’anys
Els animals que presenten una major grandària corporal requereixen també cervells més grans perquè es produeixi una coordinació adequada de les seves funcions sensorials, motores vegetatives i cognitives. Animals de grans dimensions, com elefants i balenes, posseeixen cervells molt més grans que el de l’ésser humà, malgrat la qual cosa no donen mostres d’una intel·ligència superior a la d’aquest. Això és així perquè les capacitats cognitives semblen tenir més relació amb la mida cerebral relativa que amb la mida cerebral absoluta. La mida cerebral relatiu es calcula mitjançant el quocient o índex d’encefalització, que s’obté mitjançant una fórmula logarítmica que mesura la disparitat entre la mida que hauria de tenir l’encèfal d’un animal (mida esperada) i la mida que en realitat té (mida trobada). Quan l’índex d’encefalització d’una espècie és igual a 1 vol dir que la mida esperada i trobada és igual i que per tant posseeix un cervell que correspon a la seva mida corporal. Si és menor que 1 l’espècie té un encèfal menor al que li correspondria, mentre que els índexs d’encefalització superiors a 1 indiquen la possessió de cervells més grans als que s’esperava. Els mamífers i algunes aus tenen índexs d’encefalització molt superiors als dels rèptils amfibis i peixos. Entre els mamífers, el gos, com la major part dels carnívors, té un índex d’encefalització d’un: la mida del seu cervell és l’esperat per a un mamífer del seu pes corporal. L’elefant posseeix un cervell de 5.000 grams, enfront dels 1.350 d’Homo sapiens, malgrat la qual cosa el seu índex d’encefalització és de 1,9, gairebé el doble que el d’un mamífer mitjà, però clarament inferior al de l’ésser humà.
Els primats són mamífers molt encefalitzats i els índexs de gairebé tots ells són superiors a 1. L’índex d’encefalització del ximpanzé és de 2,6 i el de l’ésser humà de 7,1, el que vol dir que el cervell de l’Homo sapiens és més de 7 vegades més gran del que li correspondria a un mamífer amb el nostre pes corporal [5].
Gran part de l’augment de la mida cerebral en les diverses espècies d’homínids sembla haver estat proporcional a l’augment de la massa corporal, però es van produir augments sense relació a l’increment de massa corporal en dues ocasions.
- La primera d’elles va tenir lloc fa aproximadament dos milions d’anys, amb les primeres espècies d’Homo: Homo habilis, Homo rudolfensis i Homo ergaster. Els Homo habilis i Homo rudolfensis van desenvolupar cervells entre 600 cc i 750 cc, enfront dels 450 cc dels Australopithecus. Homo ergaster presentava un cervell de 800 cc, però el seu índex d’encefalització no era significativament superior al d’Homo habilis / rudolfensis, atès que era proporcional a l’augment de la seva massa corporal. Encara que es va produir un augment de la mida cerebral absolut la mida cerebral relatiu no va augmentar significativament durant el milió d’anys que va seguir a l’aparició d’Homo ergaster.
- El segon increment va tenir lloc durant el Plistocè mitjà, fa entre 600.000 i 200.000 anys [6]. Aquest augment es va produir de forma independent a l’Àfrica, en els avantpassats d’Homo sapiens, i a Europa, en els avantpassats dels neandertals. La mida cerebral no ha experimentat nous augments en els últims 200.000 anys. En els últims 35.000 anys s’ha produït una disminució del 10 per 100 de la mida cerebral mitjà en la nostra espècie. Però la mida corporal de l’ésser humà va disminuir un 12 per 100 en el mateix període, pel que no es va produir una disminució de la mida cerebral relativa.
Ara bé, les troballes de Bruce Lahn [7] i col·laboradors indiquen que alguns dels gens relacionats amb l’augment de la mida cerebral semblen haver continuat evolucionant, sotmesos a la selecció natural, fins a èpoques molt recents i que potser continuïn fent-ho encara. Entre els gens que han estat sotmesos a una selecció positiva en l’ésser humà, és a dir, que han estat afavorits per la selecció natural, es troben gens relacionats amb l’augment de la mida cerebral, com ASPM i microcefalina. La variant més freqüent de microcefalina va aparèixer al voltant de 37.000 anys, i va augmentar la seva freqüència en forma extraordinàriament ràpida, el que indica que va ser seleccionada per conferir determinats avantatges adaptatius. El mateix va passar amb una variant del gen ASPM, també relacionat amb l’augment de la mida cerebral, només que, en aquest cas, la variant seleccionada va aparèixer fa uns 5.800 anys, és a dir, ja en època històrica, coincidint amb l’inici de la construcció de les primeres ciutats [8].
Atès que l’augment de mida cerebral ja havia cessat feia temps en l’ésser humà quan es van produir les mutacions, aquestes han d’haver proporcionat altres avantatges adaptatius que no coneixem encara, possiblement de tipus cognitiu, encara que no forçosament. Alguns autors suggereixen que aquests avantatges podrien estar relacionats amb un augment de l’habilitat lingüística, però existeixen encara poques dades a favor d’aquesta hipòtesi.
El cas de Flores
L’augment de grandària cerebral condueix a una reorganització estructural, amb increment de la interacció entre diverses àrees. Però ¿explica el simple augment de la mida cerebral les especialitzacions pròpies del cervell humà, com el llenguatge, la capacitat de crear i transmetre una cultura simbòlica o les capacitats de construir i utilitzar eines o representar-nos els estats mentals dels altres? L’augment de la mida cerebral, per si sol, incrementaria potser la capacitat de processar la informació i la intel·ligència general, però podria no ser capaç d’induir les especialitzacions cerebrals específiques d’una espècie. Aquestes especialitzacions es desenvoluparien únicament en el cas de produir-se canvis cerebrals associats a l’adaptació a nous entorns ecològics. En aquest cas es seleccionarien determinats gens que conduirien a canvis en la diferenciació i connectivitat neuronal i al desenvolupament de noves capacitats perceptives, conductuals i cognitives [9].
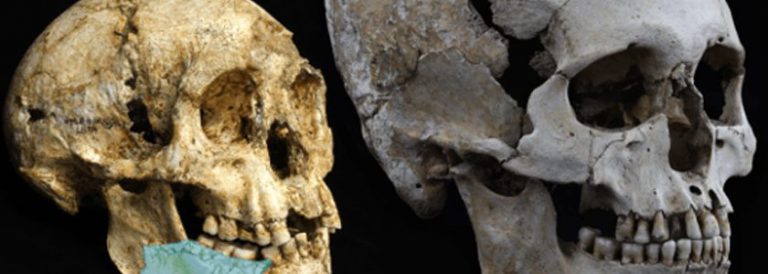
Comparació de la mida dels cranis de l’homo floresiensis i de l’homo sapiens
Esquelet del LB1 de la cova de Liang Bua, d’una antiguitat de 18.000 anys
A l’illa de Flores, a l’est de Java, s’han descobert recentment restes d’individus de només 18.000 anys d’antiguitat, l’altura dels quals no sobrepassava un metre i amb una capacitat cerebral de només 400 cc [10]. Les restes d’aquests individus estaven associades a instruments lítics i alguns paleoantropólegs opinen que podrien ser descendents d’Homo erectus, els quals haurien reduït la seva mida corporal i cerebral en adaptar-se a les condicions de vida insulars, fet que s’ha produït en nombroses ocasions en altres espècies de mamífers. Si alguna vegada arribés a demostrar que aquests homínids, amb un cervell de grandària similar al d’un ximpanzé, van ser capaços de produir instruments lítics relativament complexos, això constituiria una prova decisiva a favor que les capacitats cognitives que ens caracteritzen com a humans estan molt més relacionades amb l’organització del cervell que amb la seva mida.
La hipòtesi més probable és que les propietats que ens caracteritzen com a humans tinguin relació tant amb l’augment de la mida del cervell com amb els canvis estructurals i funcionals que van tenir lloc durant la nostra evolució cerebral.
El preu de l’encefalització
Quan parlem de l’origen d’Homo sapiens ens hem de preguntar sobre la importància del cervell en tot el que ha succeït a la nostra evolució durant els últims 150.000 anys.
Quin preu ha hagut de pagar la nostra espècie durant el procés del part de cada nou individu, pel fet d’haver aconseguit un cervell de grans dimensions?. En adquirir la postura alçada i el bipedisme, el primer problema que vam haver d’afrontar va ser donar una resposta al procés fisiològic del part. La morfologia de la pelvis dels hominins va haver de canviar per adaptar-se a la nova forma de locomoció i les modificacions van ser determinants per al procés del part experimentat per les femelles. La nostra postura bípeda va implicar no només una disminució de la mida de l’isqui i el pubis, sinó un canvi molt notable en la geometria de la pelvis i en el recorregut que ha de seguir el fetus durant el seu viatge des de l’úter matern cap a l’exterior. El canal del part va deixar de tenir una trajectòria rectilínia, per experimentar un gir molt característic. En els ximpanzés el cap del nadó és notablement més petit que el diàmetre del canal de part i durant el part es situa amb la cara mirant cap a la seva mare. Aquest fet és molt important, perquè la mare podrà netejar a la seva cria acabada de néixer amb gran facilitat. Les mares ximpanzé prefereixen la solitud de la nit per donar a llum. Es basten a si mateixes. Nosaltres hem perdut aquest avantatge.
El part en la nostra espècie és molt complex, a causa del recorregut que ha de realitzar el fetus des de la seva posició original fins que surt del ventre de la mare. El cap del nadó és molt gran pel que fa al canal del part. És el que tècnicament es denomina desproporció cefalo-pèlvica. Els girs durant el seu trànsit pel canal del part són necessaris per salvar els esculls que troba en el seu camí i per la peculiar forma del nostre maluc. El cap és massa gran per seguir una trajectòria rectilínia, com succeeix en altres primats, i ha de canviar la seva orientació a mesura que el part progressa. La posició del cap ha de posar-se de costat per adaptar-se a la forma de l’estret superior. L’amplada de les espatlles també representa un problema, que el fetus ha de solucionar en el seu recorregut des de l’estret superior fins a l’estret inferior, incloent la dificultat afegida que suposa la presència de les espines ciàtiques, que a meitat del camí produeixen l’estrenyiment del canal del part. El fetus es troba amb la necessitat de girar primer el cap i després la resta del cos per afrontar la sortida de la pelvis a través de l’estret inferior, formada per l’anell ossi que formen les tuberositats isquiàtiques, la símfisi púbica i el còccix (la part inferior de l’os sacre). La rotació del cap, però, no acaba amb la mateixa posició que en els ximpanzés, mirant cap a la mare, sinó al revés, presentant l’occipital i amb la cara cap per avall. Si la mare intervé en aquest moment les conseqüències podrien ser funestes per al seu fill, en no controlar l’esforç que hauria de fer sobre la seva columna vertebral. Davant la dificultat i el risc que comporta donar a llum a Homo sapiens, el part ha de ser assistit per la llevadora o per persones habituades a aquests menesters. En definitiva, el nostre part és un acte social, enfront de la solitud del part en altres primats.
La forma de la pelvis i del canal del part mostren el major dimorfisme sexual de tot l’esquelet humà i no és difícil esbrinar el sexe d’un individu del passat si es conserva la seva pelvis.
És possible que els nostres malucs, més estrets tinguessin una conseqüència negativa en el moment del part. Però aquest problema va poder compensar-se amb una major eficàcia en la locomoció. El menor cost energètic en carrera dels individus d’Homo sapiens va poder suposar un avantatge important sobre les altres espècies que ens van precedir. La nostra enorme resistència per córrer llargues distàncies només és possible si la despesa energètica del moviment que comporten aquests desplaçaments és raonable.
La «hipòtesi del teixit car»
D’altra banda, Leslie Aiello i Peter Wheeler [11] es van preguntar fa alguns anys sobre les raons del cost tan exagerat que comporta el creixement, desenvolupament i funcionament del nostre cervell. Ni més ni menys que entre el 20 i el 25% del metabolisme basal està dedicat a mantenir el funcionament del cervell que, com bé sabem, no descansa encara dormim profundament. El 1995 Aiello i Wheeler van proposar l’anomenada «hipòtesi del teixit car» que segueix sent acceptada per la majoria dels especialistes. Fa uns 2,5 milions d’anys la ingesta de carn es va anar incrementant en els nostres avantpassats africans. La progressiva desaparició de la cobertura vegetal que ens havia protegit durant tota la nostra trajectòria evolutiva ens va deixar a mercè de molts predadors i va fer desaparèixer alguns dels aliments bàsics de la nostra dieta. La caça d’animals petits, a la mida de les nostres possibilitats físiques, la recol·lecció d’invertebrats o l’aprofitament de la carn i el greix d’animals grans matats pels predadors de les sabanes van anar substituint una part significativa de la nostra dieta gairebé vegetariana. Com a conseqüència dels canvis en la dieta, la selecció natural va ser més «permissiva» amb la reducció i la simplificació del nostre aparell digestiu. Ja sabem que els mamífers exclusivament vegetarians necessiten un aparell digestiu llarg i complex, per digerir els hidrats de carboni de cadena llarga dels quals s’alimenten. A poc a poc, els nostres avantpassats van reduir la quantitat d’energia necessària per al desenvolupament i manteniment d’un aparell digestiu complex, que ja no necessitàvem. L’energia sobrant va poder aprofitar-se per desenvolupar i mantenir un cervell la dimensió i complexitat del qual eren cada vegada més necessaris en molts aspectes de la nostra vida, inclòs el desenvolupament d’estratègies complexes per aconseguir aliment.
Canvis en la forma del cervell
El nostre cervell és tres vegades més gran que el dels simis antropoides. La seva escorça cerebral (neocòrtex) té un gruix d’entre 2 i 5 mil·límetres, una extensió de més de 2.000 centímetres quadrats i està format per sis capes, cadascuna amb la seva peculiaritat. L’escorça està formada per multitud de columnes o «mòduls» independents entre si, però connectats a través de l’anomenada capa IV. Finalment, cal destacar que l’escorça no és uniforme al llarg de tot el cervell, sinó que la seva composició és variable en relació amb el seu paper en el funcionament cerebral.
Especialistes com Philip Gunz i Simon Neubauer [12] han trobat una diferència important entre la nostra espècie i els altres hominins pel que fa al cervell. Aquests investigadors han estudiat la forma del cervell dels ximpanzés, els neandertals i els humans moderns. El mètode emprat (morfometria geomètrica) elimina el factor mida i considera només les variacions de la forma que succeeixen durant el desenvolupament del cervell des del naixement fins que aquest òrgan arriba a la seva mida definitiva. Recordem que els neandertals van arribar a tenir mides cerebrals superiors als d’Homo sapiens, mentre que els humans del Plistocè Mitjà no van estar lluny d’arribar a una mida cerebral com el nostre. Günz i Neubauer han observat que la trajectòria ontogenètica de la forma del cervell humà és molt similar a la dels ximpanzés i a la dels hominins, excepte durant el primer any de vida. Aquí rau la gran diferència, que reflecteix un model ben definit fa alguns anys per Daniel Lieberman [13] i els seus col·laboradors. El neurocrani experimenta una «fase de globularització», en la qual el crani adopta una forma notablement esfèrica. Recordem els cranis allargats i més o menys baixos d’Homo erectus i Homo neanderthalensis per adonar-nos de les diferències amb el crani més curt i alt d’Homo sapiens. La globularització es produeix mitjançant una expansió relativa del lòbul frontal, dels lòbuls temporals i de la fossa cranial posterior, un aplanament de l’àrea occipital i una flexió de la base del crani. Els lòbuls temporals semblen començar la seva expansió en altres espècies d’hominins, de manera que l’amplada màxima del crani es desplaça cap amunt, en lloc de situar a la part més baixa del crani. Sembla, doncs, clar que Homo sapiens va donar el salt cap a una trajectòria ontogenètica diferent del cervell. El cerebel d’Homo sapiens és més gran del que s’esperava per a un primat de la nostra mida corporal.
Finalment, la globularització cerebral implica una notable flexió de la base del crani. La flexió de la base del crani a Homo sapiens es produeix en un moment en què la velocitat de creixement del cervell és molt elevada. La fase de globularització del primer any de vida sembla ser única de la nostra espècie i, per tant, un tret derivat que no està present ni en els ximpanzés ni en altres espècies d’hominins de la nostra genealogia. És per això que ens podem preguntar: ¿tenim capacitats intel·lectuals superiors gràcies a aquest canvi en el desenvolupament de la forma del cervell?
L’altricialitat secundària
El desenvolupament del cervell dels vertebrats comença amb el naixement de les cèl·lules progenitores de les futures neurones. Segueix amb un procés de diferenciació cel·lular i una migració de les cèl·lules cap a la seva ubicació definitiva al cervell. El creixement de dendrites i axons produeix una complexa trama de connexions entre les diferents cèl·lules cerebrals mitjançant milions de sinapsis formant una xarxa, que de moment els experts són incapaços de desxifrar. El pas següent consisteix en la «poda» de moltes d’aquestes sinapsis que van ser utilitzades durant la fase embrionària. Finalment el procés es completa amb la protecció dels axons mitjançant una beina de mielina, que multiplica per cent la velocitat de transmissió dels impulsos nerviosos. La complexa regulació genètica del programa que controla tot el procés no es coneix encara. Cada espècie té el seu propi programa que finalment determina la mida, la forma de cadascuna de les regions cerebrals i de tot el cervell. així com la complexitat de la trama de connexions neuronals. En aquest procés, la velocitat, el temps i el moment relatiu de l’inici i final de cada esdeveniment tenen un paper fonamental en el desenvolupament cerebral. Petits canvis en aquests factors (heterocronies del desenvolupament) poden tenir conseqüències de gran calat.
Una qüestió fonamental del nostre desenvolupament cerebral és l’anomenada altricialitat secundària, un terme introduït per Adolf Portmann en 1969. Naixem amb un enorme retard neuromotriu i cognitiu respecte a altres mamífers. Quan naixem, tot just ens movem i emetem un plor dèbil per mostrar fam, fred o dolor. Encara trigarem com a mínim un any per donar els nostres primers passos com a primats bípedes. Per contra, els ximpanzés corren en pocs mesos per la selva.
Hipòtesi del «dilema obstètric»
Portmann va proposar la hipòtesi del «dilema obstètric» per explicar aquesta peculiaritat humana. La bipedestació va reduir les dimensions del canal del part fins a un punt crític, com hem vist. Ja que la nostra taxa de creixement cerebral és molt elevada durant la gestació hem de néixer abans d’haver completat el desenvolupament òptim de les funcions cerebrals. Tots els simis antropoides necessiten de la cura de les seves mares, però el temps de dependència és molt més curt que en la nostra espècie. Amb l’excepció del temps de lactància, els petits simis es mouen amb gran agilitat pel seu mitjà natural. Segons Portmann, la nostra gestació hauria de durar uns 18-21 mesos per néixer en les mateixes condicions neuromotores que els nostres cosins els simis antropoides.
«Hipòtesi metabòlica»
El 2012, Ral·li Dunsworth [14] i altres autors van proposar una hipòtesi diferent, que van denominar «hipòtesi metabòlica». Segons aquests investigadors, el senyal per a l’inici del part està governada pel balanç entre el metabolisme de la mare i el del fetus, mitjançant una determinada concentració hormonal. Els éssers humans tenim una taxa metabòlica basal, condicionada precisament per la quantitat d’energia que necessita el cervell. Podem augmentar la despesa energètica en situacions límit, multiplicant fins per quatre o cinc vegades aquesta taxa basal. Això pot succeir durant lapses breus de temps. Ens pot valer l’exemple d’un ciclista pujant un port de primera categoria a ritme infernal: el ciclista arribarà exhaust després de l’esforç suprem, ara bé, si aquest esforç dura molt de temps l’esportista pot arribar al col·lapse. Una mare gestant arriba a valors de fins a 2,5 vegades la seva taxa metabòlica basal. Però el temps dedicat a aquest esforç és continuat. La mare necessita energia per mantenir el seu propi cos i la seva activitat normal, al que cal sumar el desenvolupament del fetus i dels teixits que el suporten, com la placenta. No oblidem que el cervell del fetus està creixent a gran velocitat. Arribat un moment, la mare és incapaç de gastar més energia i és llavors quan es produeix el senyal per al part. La lactància seguiria sent una exigència energètica molt important per a la mare, però la despesa total per tirar endavant la cria es repartirà en el temps, sense necessitat d’arribar a valors extrems.
Les dues hipòtesis no tenen per què ser mútuament excloents. Més aviat al contrari, podrien sumar els seus efectes perquè es produís el senyal per al part.
El procés de mielinització
D’altra banda, en les neurones la formació de la beina protectora de mielina té un paper crucial en l’activitat dels axons. La beina de mielina no és contínua, sinó que hi ha diminutes interrupcions d’un micròmetre de longitud anomenades nòduls o nodes de Ranvier. La presència d’aquests nòduls fa que l’impuls nerviós viatgi «a salts», amb una velocitat notablement més elevada a la dels axons que no tenen mielina i sense possibilitat d’error en la seva direcció. El color blanquinós de la mielina dóna nom a la substància blanca del neocòrtex, mentre que l’absència d’aquesta substància (per exemple, en els cossos neuronals) dóna lloc al que anomenem «substància grisa». Se sap que la densitat de fibres mielinitzades està relacionada amb la maduresa de les diferents àrees cerebrals. Com més gran sigui aquesta densitat, major serà la maduresa de les regions cerebrals i menor la seva capacitat de respondre a nous estímuls del medi ambient. Segons els estudis de Daniel J. Miller i un nombrós equip de col·laboradors [15] centrat en el neo-còrtex de les àrees sensorial, motora, visual i prefrontal, els nounats humans no tenim mielina en el moment del naixement, mentre que el 20% d’aquestes àrees ja està mielinitzada en els ximpanzés al final del procés de gestació. En aquests primats la mielinització acaba durant la maduresa sexual (aproximadament cap als 11 anys) en la major part del neocòrtex cerebral, excepte en l’àrea frontopolar. Aquesta àrea, on resideixen funcions com la planificació, la presa de decisions o l’anticipació dels esdeveniments, acaba el seu procés de mielinització cap als 17 anys. Fins a aquesta edat, l’àrea frontopolar dels ximpanzés encara pot enriquir-se amb nous estímuls.
Pel que fa als éssers humans, Miller i els seus col·laboradors han observat un retard considerable del procés de mielinització en totes les àrees del neocòrtex cerebral, que finalitza a prop dels 30 anys, una edat que pot resultar sorprenent per a alguns lectors. Com succeeix en els ximpanzés, el major retard es produeix en l’àrea frontopolar, en què la poda de sinapsis utilitzades durant el primer terç de la vida es retarda de manera espectacular i única entre els primats. D’altra banda, James K. Rilling ha demostrat mitjançant l’estudi d’imatges obtingudes per ressonància magnètica que les nostres àrees cerebrals associatives, com la mateixa àrea prefrontal, tenen una menor densitat de mielina que les àrees motores i sensorials. Mentre que aquestes últimes arriben a la seva plena maduresa, les àrees associatives del neocòrtex del nostre cervell romanen amb certa capacitat d’adaptació durant molts anys. En altres paraules, els humans actuals disposem d’una alta capacitat conceptual i d’abstracció durant la major part de la nostra vida respecte a altres primats, com els ximpanzés o els macacos. No obstant això, com han demostrat altres investigadors, el cost energètic dels canvis experimentats per Homo sapiens no és menyspreable. La nostra plasticitat cerebral, basada en una menor densitat de mielina en certes regions i en la possibilitat de formació de sinapsis durant la vida adulta, implica una despesa energètica molt superior al d’altres primats. A més, el retard en la maduresa cerebral pot suposar l’aparició de malalties com l’esquizofrènia, o produir el perill físic que suposa la immaduresa dels adolescents durant un llarg període de temps. És el preu que hem de pagar per haver aconseguit capacitats cognitives úniques entre els primats vius.
Reorganització cerebral
Hi ha nombroses evidències anatòmiques que en l’evolució cerebral humana s’ha produït no només un augment de grandària, sinó una reorganització estructural, que va fer possible el desenvolupament de les capacitats cognitives que caracteritzen la nostra espècie.
- Fa entre 3 i 2 milions d’anys es va produir un augment de mida del lòbul frontal dret respecte l’esquerre i de la zona parietooccipital esquerra pel que fa a la dreta.
- Entre 2,5 i 1,8 milions d’anys es va produir una reorganització del lòbul frontal en les primeres espècies del gènere Homo, sobretot de la tercera circumvolució frontal inferior, la qual conté l’àrea de Broca, relacionada amb la producció del llenguatge, que va adquirir un aspecte semblant al que presenta en l’ésser humà actual.
- En les primeres espècies d’Homo també es va produir una reducció del còrtex visual primari.
- En els últims 500.000 anys es va produir un augment de la mida cerebral, però els motlles endocranials no revelen canvis significatius en la morfologia cerebral frontal durant aquest període [16].
Els estudis de Bruner amb motlles endocranials, però, demostren que el cervell dels éssers humans anatòmicament moderns es caracteritza, sobretot, per l’augment de mida dels lòbuls parietals. L’augment de mida parietal sembla haver-se iniciat ja en els Australopithecus molt abans que es produís l’augment de mida dels lòbuls frontals. Els lòbuls parietals són àrees d’associació que intervenen en la integració de diferents modalitats sensorials, en la coordinació espacial, en l’atenció i en la representació interna de la realitat. El sistema frontoparietal a Homo sapiens constitueix una complexa xarxa relacionada amb les facultats mentals humanes més elevades, com la capacitat de raonar [17].
A partir de finals del Plistocè i durant l’Holocè sembla haver-se produït un lleuger augment de mida del cerebel en relació als hemisferis cerebrals. El cerebel regula la coordinació motora, presenta múltiples connexions amb el neocòrtex i intervé en la seqüenciació motora complexa i en alguns aspectes cognitius, com l’habilitat lingüística, pel que el seu major desenvolupament en l’Holocè i últimes fases del Plistocè pot haver resultat adaptatiu en un món de creixent complexitat cultural [18].
No totes les regions cerebrals van augmentar en mida o complexitat durant l’evolució del gènere Homo. Gran part de l’augment de la mida cerebral va ocórrer gràcies a l’expansió del neocòrtex. La proporció entre la substància grisa del neocòrtex i el tronc cerebral és de 30: 1 en els ximpanzés i de 60: 1 en els éssers humans moderns. En relació als grans simis, en els éssers humans s’ha produït no només un augment global del neocòrtex, sinó també un major nombre de circumvolucions a les esperades en les àrees d’associació, la qual cosa implica la possessió d’unes àrees d’associació relativament majors, i un augment de la substància blanca en relació a la substància grisa cerebral, particularment evident en la substància blanca prefrontal, desproporcionadament més gran en els éssers humans que en els primats no humans [19]. L’augment de mida de les àrees d’associació reflecteix l’increment de la capacitat de processament cerebral en aquestes àrees. L’augment relatiu de la substància blanca indica un major nombre de connexions entre neurones neocorticals.
En l’ésser humà s’observen asimetries anatòmiques entre ambdós hemisferis cerebrals, que reflecteixen una lateralització i especialització hemisfèrica en diferents funcions o dominis. La major part de funcions relacionades amb el llenguatge es duen a terme a l’hemisferi esquerre, encara que altres, com la prosòdia o entonació emocional, tenen lloc a l’hemisferi dret. Com en la pràctica totalitat de funcions cognitives, en el llenguatge hi participen diverses regions cerebrals interconnectades. Les funcions cognitives no solen tenir lloc en àrees aïllades, sinó en xarxes que comprenen diferents zones, corticals i subcorticals. Una mateixa regió cerebral pot participar en diferents funcions cognitives. En el llenguatge intervenen no només les àrees de Broca i Wernicke, situades respectivament en els lòbuls frontal i temporal esquerres, sinó altres àrees adjacents, com el lòbul prefrontal, i estructures subcorticals, com els ganglis basals, els quals participen també en l’execució de moviments seqüencials complexos.
La utilització d’eines activa el còrtex esquerre, sigui quina sigui la mà utilitzada. Les evidències neurofisiològiques indiquen que l’ésser humà presenta especialitzacions cerebrals, tant per a representar les interaccions dels objectes físics com per a la representació de les accions manuals relacionades amb l’ús d’eines. Aquestes funcions estan representades en el còrtex temporal superior i el còrtex parietal posterior i el còrtex premotor, respectivament [20]. Mitjançant noves tècniques de neuroimatge, com la ressonància magnètica DTI, s’ha detectat en l’ésser humà un feix de fibres que connecten el còrtex temporal mitjà amb el còrtex parietal inferior. Aquest feix de fibres, molt més gruixut en l’hemisferi esquerre, és gairebé inexistent en els grans simis i podria tractar-se d’una especialització del cervell humà relacionada amb la utilització d’eines [21].
L’hemisferi dret en els humans porta a terme les funcions d’atenció i visuospaciales. El reconeixement de rostres, de gran importància adaptativa en la nostra espècie, té lloc bilateralment a la cara inferior de la regió tèmporo-occipital. Des del punt de vista anatòmic, en el cervell humà s’observa un augment de mida del lòbul frontal dret respecte l’esquerre i de la regió parietooccipital esquerra pel que fa a la dreta. La fissura de Silvio presenta una major longitud en l’hemisferi esquerre i la regió denominada planum temporale és més gran en l’hemisferi esquerre que en el dret. Aquesta regió comprèn part de l’àrea de Wernicke implicada en la comprensió del llenguatge.
Aquestes diferències anatòmiques van ser interpretades inicialment com la demostració que l’ésser humà presentava una lateralització molt més gran que la resta dels primats. Estudis més detallats van demostrar posteriorment que les asimetries en la fissura de Silvio s’observaven també en els primats no humans. S’ha demostrat recentment que els ximpanzés presenten asimetries del planum temporale similars a les humanes i que, en els grans simis, l’àrea homòloga a la de Broca també és més gran en l’hemisferi esquerre. Mitjançant ressonància magnètica DTI, s’ha descobert recentment que el fascicle arquejat esquerre humà que connecta les àrees de Broca i Wernicke, que intervenen respectivament en la producció i comprensió de llenguatge és molt més gruixut del que s’havia cregut fins fa poc i que les seves terminacions posteriors no es limiten a l’àrea de Wernicke, sinó que s’estenen per una gran part del còrtex temporal mig i inferior, una adaptació humana relacionada sens dubte amb la capacitat de llenguatge (Rilling, 2008).
En general, es pot afirmar que l’ésser humà posseeix les mateixes àrees neocorticals que els grans simis encara que algunes d’aquestes àrees i les connexions que hi ha entre elles han experimentat importants variacions de mida en la nostra espècie. Algunes regions del cervell humà no només no van experimentar augments de mida i complexitat sinó que es van tornar més petites i simples, com el bulb olfactori. L’àrea visual primària humana presenta una grandària menor a l’esperat per a un neocòrtex com el nostre: aquesta àrea s’ocupa un 5 per 100 del còrtex del ximpanzé i només un 2 per 100 en el còrtex humà. La zona del lòbul temporal humà relacionada amb el processament del llenguatge i altres estímuls auditius és més gran al que s’espera per un primat amb la nostra mida cerebral.
Els experts en neuroanatomia comparada han vingut sostenint des de fa anys que el més característic de l’evolució del cervell humà va ser l’augment de mida dels lòbuls frontals. Aquesta afirmació va ser refutada per un estudi dut a terme per Semendeferi i col·laboradors, en el qual es van comparar, mitjançant ressonància magnètica, els cervells d’humans, simis i micos. Les conclusions de l’estudi van ser que el lòbul frontal humà, que ocupa un 35 per 100 del volum cortical total, no era significativament més gran que el dels grans simis, en termes relatius[22]. En termes absoluts, però, el lòbul frontal humà és molt més gran que el de qualsevol simi. Mentre que les àrees motores primàries del lòbul frontal no han augmentat de grandària en el cervell humà en termes absoluts, sí hi ha hagut un augment significatiu de mida, absolut i relatiu, en les àrees d’associació. El còrtex prefrontal lateral ocupa un 29 per 100 del neocòrtex humà i només un 17 per 100 del d’un ximpanzé. Particularment l’àrea 10 del còrtex prefrontal humà és més gran, en relació a la resta del cervell, que la seva homòloga en els grans simis i presenta un major nombre de connexions amb altres àrees d’associació [23].
El còrtex prefrontal lateral està interconnectat amb altres àrees que han augmentat de mida durant l’evolució del cervell humà, com a parts del còrtex temporal i el còrtex parietal posterior. Sembla que el còrtex prefrontal lateral humà exerceix un major control que a la resta dels primats sobre altres àrees cerebrals. El còrtex prefrontal lateral intervé en funcions com l’evocació de la memòria episòdica, la memòria operativa, el control del llenguatge i la planificació de la conducta. Basant-se en coneixements i records, intervé en la capacitat d’imaginar diversos escenaris possibles, trobar analogies, representar noves solucions per resoldre problemes i en la realització d’actes voluntaris, planificant, controlant i suprimint diversos actes motors. L’augment de grandària i complexitat del còrtex prefrontal lateral, va permetre a l’ésser humà una major flexibilitat i varietat en la conducta, la capacitat d’imaginar, fabricar i utilitzar instruments i l’habilitat de representar-se els objectes simbòlicament, sense la qual no seria possible el llenguatge.
El cerebel
El cerebel de la nostra espècie té, com passa amb el cervell, una mida superior a l’esperat per un primat de la nostra mida corporal en relació a altres espècies, com els macacos, els micos caputxins, els babuïns o els ximpanzés. De fet, el cerebel ocupa el segon lloc en el rànquing de les parts cerebrals que més han augmentat respecte a la nostra mida corporal, per darrere del neocòrtex cerebral. El cerebel s’ha considerat de manera tradicional com una part del cervell implicada en funcions motores. No obstant això, les investigacions anatòmiques i neurofisiològiques més recents sobre el cerebel mostren el seu paper en la cognició. Segons alguns investigadors, el cerebel podria estar relacionat amb la velocitat d’altres funcions del neocòrtex. La connexió entre el cerebel i el neocòrtex permetria una major velocitat i possibilitats tant en les funcions motores com en les capacitats cognitives.
El lòbul frontal del neocòrtex
Per descomptat, el lòbul frontal del neocòrtex té molt a veure amb el gran desenvolupament de certes capacitats cognitives en la nostra espècie. Aquesta regió té tres elements fonamentals: el còrtex prefrontal dorsolateral, que representa la regió més anterior del cervell, el còrtex orbital prefrontal, que només es podria veure des de la part inferior del cervell, i el còrtex prefrontal medial, que podríem observar tallant sagitalment el cervell. En Homo sapiens aquestes regions cerebrals han augmentat la seva mida en major mesura que les altres, en comparar la seva mida amb la d’altres primats.
- El còrtex prefrontal dorsolateral és bàsic en una cosa tan fonamental com l’anticipació, la planificació i la presa de decisions, que podria ser una manera de definir la intel·ligència. Els éssers humans acumulem (conscient i inconscientment) una enorme quantitat d’informació sobre fets ocorreguts al llarg de la nostra vida, de manera que podem anticipar-nos a les circumstàncies i solucionar problemes sobrevinguts mitjançant l’ús adequat d’aquesta informació.
- El còrtex orbital prefrontal té un paper determinant com a mediador entre les emocions i els pensaments.
- El còrtex prefrontal dorsolateral s’ha relacionat en part amb l’anomenada «teoria de la ment», o la capacitat de representació dels estats mentals propis i els dels altres, el que ens porta a atrevir-nos a pensar el que altres creuen , desitgen o pensen.
Còrtex cingular anterior
En parlar de la nostra capacitat intel·lectual és fonamental tenir en compte l’important paper del cíngol anterior, que se situa en el còrtex prefrontal medial i que representa un intermediari entre les emocions i vivències i les solucions aportades en cada moment de la vida. Està implicat en funcions bàsiques com la pressió arterial, el funcionament correcte de la digestió i la taxa cardíaca. A més, també participa activament en la regulació de processos cognitius de certa complexitat, relacionats amb l’avaluació entre els problemes que ens imposa el medi ambient i la solució que adoptem en cada cas. En aquest procés és fonamental l’amígdala potser l’element més important del sistema límbic per rebre les emocions que ens arriben cada dia i que confrontem amb el còrtex prefrontal. Tot això pot explicar la raó per la qual les emocions experimentades ens puguin jugar una mala passada durant la digestió o ens elevin puntualment la pressió arterial. Molts dels nostres presumptes problemes de salut deixen d’existir, com per art de màgia, en distanciar-nos per un moment del fort estrès del món civilitzat i dels seus conseqüents problemes emocionals.
Canvis en el desenvolupament cerebral
En adquirir la posició bípeda es va produir un estrenyiment de la pelvis i, per tant, del canal del part. Els primers membres del gènere Homo, en presentar un cervell més gran, precisaven canals del part més amples, el que implicava una pelvis també més ampla. Però una correcta marxa bípeda imposava un límit a l’amplada pèlvica femenina. La solució trobada per la selecció natural va ser que les cries naixessin en una etapa més immadura. Encara que en l’ésser humà el període de gestació és lleugerament més llarg que l’esperat per un primat de la seva grandària, aquest neix en una fase més immadura que la resta dels primats; d’aquesta manera, gran part de l’increment del desenvolupament cerebral es produeix després del naixement. Posposant una gran part del desenvolupament cerebral al període postnatal, els avantpassats de l’ésser humà van poder augmentar la seva mida cerebral sense un excessiu augment de la mida cranial del nadó. Els grans simis però neixen també en fases relativament immadures pel que aquesta tendència sembla haver-se iniciat en etapes anteriors al començament del bipedisme. En néixer, el cervell d’un macaco ha completat ja el 70 per 100 del seu desenvolupament, mentre que el d’un ximpanzé només ha completat un 40 per 100 i el d’un ésser humà únicament un 25 per 100 [24]. El cervell dels simis antropoides i especialment l’humà, està, per tant, exposat a un major nombre d’estímuls durant la seva etapa de desenvolupament, el que potser podria facilitar la producció d’un major nombre de connexions sinàptiques.
El cervell humà presenta també un desenvolupament molt més lent que el dels primats no humans. Tant macacos com a éssers humans inicien la mielinització cerebral abans del naixement. La mielinització consisteix en el recobriment dels axons, o prolongacions neuronals, per una capa de proteïnes i lípids anomenada mielina. La substància blanca cerebral està constituïda majoritàriament per mielina. Aquesta capa de mielina permet una millor conducció dels impulsos nerviosos millorant les connexions entre diferents àrees cerebrals i el funcionament cerebral global. Ambdues espècies mostren un patró de mielinització similar en el moment del naixement excepte pel que fa a la mielinització del neocòrtex present ja en macacos nounats però no en humans en els que s’inicia un mes després del naixement i continua fins als vuit anys. En els macacos la mielinització cortical es produeix entre tres i quatre vegades més ràpid que en els humans i acaba als dos anys d’edat [25]. El desenvolupament d’algunes àrees d’associació cerebral, especialment els lòbuls prefrontals, pot prolongar-se en l’ésser humà fins ben entrada la tercera dècada de la vida. El ritme de formació de sinapsis varia entre diferents àrees corticals: la sinaptogènesi, o formació de noves connexions sinàptiques, triga molt més a completar-se en el còrtex frontal que en l’escorça visual primària.
El període de creixement cerebral és molt més llarg en éssers humans que en els grans simis. Mentre que en el ximpanzé el cervell arriba a la mida adult als cinc anys, a Homo sapiens el període de creixement cerebral és tres vegades més gran [26]. La ràpida expansió de la mida cerebral en el gènere Homo sembla haver-se aconseguit mitjançant l’expansió del període de creixement fetal i, sobretot, infantil i el manteniment d’una taxa de creixement fetal durant un període més llarg que el dels simis antropoides, els seus parents més propers. Aquests canvis impliquen la retenció de característiques infantils durant llargs períodes de temps. A la retenció de característiques infantils a la edat adulta o durant llargs períodes se li dóna el nom de neotènia. La gran grandària cerebral humà i el seu elevat nombre de connexions semblen ser, almenys en part, el resultat de la retenció en la nostra espècie de característiques neotèniques.
Els animals de cervell gran necessiten més temps per desenvolupar-se, per la qual cosa depenen de la cura parental durant períodes molt llargs, tendència que es va accentuar significativament en el gènere Homo. Tant en els grans simis com en les diverses espècies d’Homo que han existit, els pares havien de viure molt més enllà de la maduresa sexual per assegurar la supervivència de les cries [27]. En ximpanzés, l’experiència materna incrementa l’èxit de les cries adultes i les mares contribueixen a l’estatus social dels seus fills. La selecció natural, per tant, va afavorir a aquells gens que potenciaven la longevitat dels adults. En la major part dels primats són les femelles les que tenen cura de les seves cries. Gran part de l’aprenentatge en els éssers humans depèn de la transmissió de coneixements mitjançant ensenyament intencional a través del llenguatge, és a dir, de la cultura simbòlica. En el cas particular de l’ésser humà, aquest ensenyament no es troba a càrrec exclusivament de la mare, sinó també de pares avis, oncles i altres adults.
Els estudis de Hawkes mostren que en societats de caçadors recol·lectors les dones en edat posreproductiva contribueixen a la supervivència de nebots i nets, en aportar aliments rics en calories, a més de coneixements i experiència [28]. És per això que en el cas únic de l’ésser humà, podria haver-hi hagut una selecció per a una major longevitat, superior a la que es va produir en grans simi i altres animals de gran cervell, actuant contra gens l’acció deletèria dels quals no se sol manifestar fins a edats avançades de la vida.
La longevitat humana va créixer en més d’un 20 per 100 en els últims 30.000 anys. Fins llavors la longevitat màxima s’havia situat en quaranta-cinc anys aproximadament. A la fi del Plistocè des l’inici de l’etapa cultural coneguda com Paleolític superior, comencen a ser relativament freqüents les restes d’individus d’edat superior als cinquanta anys, i fins i tot uns pocs van començar a sobrepassar els seixanta [29]. És possible que l’augment de longevitat humana, esdevingut durant el Paleolític superior, no fos degut únicament a factors culturals. La selecció de la longevitat va poder tenir relació amb la creixent importància de les relacions entre diferents generacions que va suposar l’augment de complexitat de la cultura simbòlica la qual requeria la transmissió social i emmagatzematge en la memòria de gran quantitat d’informació. En aquestes condicions, l’experiència de la gent gran va poder arribar a ser més necessària que mai [30].
Canvis genètics
El cervell humà és un òrgan extremadament complex, que porta a terme gran nombre de funcions, algunes exclusives de la nostra espècie, i en el qual s’expressen al voltant d’una tercera part dels gens que componen el nostre genoma. Ja hem vist el cas dels ASPM i les microcefalines. ¿S’ha donat una selecció positiva en altres gens relacionats amb la funció cerebral? Els avenços de la genètica i proteòmica i la recent seqüenciació dels genomes de l’ésser humà i del ximpanzé estan permetent, per primera vegada, detectar les variacions en l’estructura i expressió genètica cerebral relacionades amb els canvis estructurals i funcionals ocorreguts durant l’evolució del cervell humà.
Entre els gens que han incrementat la seva expressió en el cervell humà, la major part, com CaMK2, es troben relacionats amb la transmissió i la plasticitat sinàptica. També s’ha trobat incremento.de l’expressió de gens relacionats amb la neuroprotecció, especialment davant factors relacionats amb l’edat avançada com l’estrès oxidatiu, i de gens relacionats amb el metabolisme aerobi i la funció mitocondrial (Càceres et al., 2003; Uddin et al., 2004). Excepte els relacionats amb la neuroprotecció, la resta dels gens que havien incrementat la seva expressió en el cervell humà estaven directament o indirectament relacionats amb l’activitat i plasticitat sinàptiques. L’alt consum metabòlic de les neurones neocorticals humanes és necessari per mantenir el gran nombre de prolongacions neuronals i connexions sinàptiques que caracteritzen a aquestes neurones en la nostra espècie (Sherwood et al., 2006). Però un elevat metabolisme genera també més radicals lliures, els quals poden fer mal a la cèl·lula accelerant el seu envelliment, de manera que en l’ésser humà, més que en cap altra espècie, ha estat necessària la selecció de gens amb funció neuroprotectora.
La regió del genoma humà que ha experimentat una major acceleració evolutiva des que es va produir la separació dels llinatges que van portar als éssers humans i els ximpanzés, forma part d’un gen anomenat Harif [31] (Pollard et al., 2006). El gen Harif ha experimentat 18 substitucions en els 5 a 7 milions d’anys transcorreguts des que es van separar el llinatge humà i el del ximpanzé. En els 300 milions d’anys anteriors només havien canviat dues bases la qual cosa és la diferència observada entre el ximpanzé i les aus. Hi va haver, probablement, una forta pressió selectiva que va promoure l’evolució d’aquest gen i que només es va donar en els avantpassats de l’ésser humà. Hi ha indicis que el gen Harif influeix en l’expressió cerebral d’una proteïna de la matriu extracel·lular, anomenada reelina. La reelina intervé en el desenvolupament cerebral durant l’etapa fetal, en la qual regula la migració i el posicionament de les neurones. La seva expressió persisteix, però en el cervell adult, on modula l’activitat dels receptors NMDA, relacionats amb l’aprenentatge i intervé en la neuroplasticitat i formació de noves sinapsis interaccionant amb la proteïna ApoE amb la qual comparteix el mateix receptor neuronal [32].
La reelina sembla haver exercit un paper molt important en l’evolució cerebral. En els mamífers es va produir un increment de la síntesi d’aquesta proteïna, el que va contribuir a l’expansió del neocòrtex, en permetre l’emigració neuronal a capes més superficials durant el desenvolupament cerebral. En els primats la reelina s’expressa en un major nombre de cèl·lules cerebrals que a la resta dels mamífers. La major part dels mamífers adults expressen la proteïna només en cert tipus d’interneurones productores del neurotransmissor anomenat Gaba, mentre que el primats l’expressen també en les neurones piramidals de l’hipocamp i neocòrtex [33]. L’expressió de reelina sembla haver-se incrementat en relació directa amb l’augment de complexitat cerebral i l’increment de les capacitats de memòria i aprenentatge, les quals impliquen un augment de la neuroplasticitat. La ràpida evolució del gen Harif suggereix que poden haver-se produït importants canvis en la expressió de reelina durant l’evolució cerebral del gènere Homo. Aquests canvis poden haver tingut relació amb l’augment de la mida cerebral i de la neuroplasticitat en els adults de la nostra espècie.
Recentment s’ha detectat en el neocòrtex humà, però no en altres estructures cerebrals, com el cerebel, l’increment d’expressió de dues variants de gens que codifiquen una proteïna de la matriu extracel·lular, la trombospondina. Una de les variants (trombospondina 2) presenta una expressió dues vegades superior a la dels primats no humans, incloent el ximpanzé, i l’expressió de l’altra variant (trombospondina 4) és sis vegades superior a la dels primats no humans. Aquestes proteïnes intervenen també en la neuroplasticitat, el que constitueix una dada més a favor que en l’ésser humà s’ha produït un augment de la plasticitat sinàptica, és a dir, de la capacitat de creixement de les prolongacions neuronals o neurites i de formar noves connexions neuronals en l’adult [34].
Desconeixem encara la major part dels canvis genètics relacionats amb l’evolució cerebral en el gènere Homo. Les dades de què disposem fins al moment suggereixen que gran part d’aquests canvis constitueixen adaptacions per mantenir alts nivells d’activitat neuronal durant una llarga vida. Una gran part d’aquesta activitat neuronal consisteix en una major activitat i plasticitat sinàptiques, el que comporta un increment de les capacitats d’aprenentatge, memòria i processament de la informació. Això sembla haver-se aconseguit mitjançant relativament poques modificacions no en gens que codifiquen proteïnes sinó en gens reguladors, els quals regulen l’activitat de diferents gens estructurals o codificadors de proteïnes. Aquestes modificacions han portat a la persistència de caràcters juvenils en cèl·lules pertanyents a àrees cerebrals associades a funcions cognitives complexes.
Pressions selectives en l’evolució de la ment
Les dades aportades per les neurociències i altres disciplines estan permetent conèixer els canvis ocorreguts durant l’evolució cerebral humana, però no expliquen per què van ocórrer aquests canvis. Quines pressions selectives els van propiciar? Si els caràcters derivats del cervell humà són un reflex del medi ancestral en el qual va evolucionar el gènere Homo, la reconstrucció de les condicions de vida dels nostres avantpassats durant el Plistocè potser podria aportar algunes respostes. Sense descartar l’existència de canvis cerebrals des de la invenció de l’agricultura, fa 10.000 anys, probablement poc significatius donat el curt temps transcorregut, una gran part de l’evolució cerebral que va convertir a un Australopithecus a Homo sapiens va tenir lloc com a resposta a les pressions selectives (ecològiques, tècniques, socials i culturals) que es van donar en els últims 3 milions d’anys.
- Adaptació als canvis climàtics
El Plistocè, el període geològic en què va evolucionar el gènere Homo es va caracteritzar per importants, i de vegades molt ràpides, oscil·lacions climàtiques. S’hi van donar períodes molt secs i altres humits, períodes glacials, en què el nord d’Europa i Amèrica estaven coberts per grans masses de gel, i períodes càlids. A l’Àfrica, el nostre lloc d’origen, les enormes selves tropicals van deixar pas a les grans sabanes, caracteritzades per ser espais on els recursos es donen dispersos i de manera irregular, tant en el temps (estacions), com en l’espai (recursos diferents en llocs diversos) [35]. Els homínids van haver d’adaptar-se a aquesta heterogeneïtat i només van poder fer-ho a través de dues vies: una dieta capaç de recuperar energia de gairebé tot tipus de recursos vegetals i animals, i una cultura més complexa, enfocada entre altres coses a la plasticitat conductual i al sorgiment i perfeccionament tecnològics.
Com adaptació a aquest nou mitjà els homínids van passar a dependre en forma creixent de la carn com a font d’aliment, van començar a fabricar eines per procurar-se-la i van formar grups socials més interactius i complexos, en els quals va anar augmentant la importància de la cooperació i la comunicació, sense perdre per això els impulsos competitius i jeràrquics propis dels nostres avantpassats primats. L’augment de grandària cerebral en els primers Homo, fa més de 2 milions d’anys, coincideix amb l’augment d’ingesta de carn i la construcció dels primers instruments lítics. La ingesta de carn va permetre el creixement cerebral en els primers Homo, al mateix temps que es produïa una reducció del tub digestiu. Aquesta reducció va permetre utilitzar més quantitat de l’energia procedent dels aliments per construir un cervell més gran i més complex[36]. Però el cervell és un òrgan energèticament car: encara que el seu pes representa únicament el 2,3 per 100 del pes total del cos, consumeix més del 20 per 100 de l’energia utilitzada per l’organisme. El simple fet de menjar carn no hagués produït un augment espectacular de la grandària cerebral si aquest augment no hagués representat una important avantatge selectiu, que va contribuir a augmentar les possibilitats de supervivència i de produir descendència viable en els nostres avantpassats.
La dependència de l’aportació de carn va anar augmentant progressivament en les primeres espècies d’Homo. Però, a diferència d’altres depredadors, com felins i cànids, els homínids no havien evolucionat com depredadors. Aquests han coevolucionat amb les seves preses al llarg de milions d’anys, en una «carrera d’armaments» en què caçadors i preses van anar desenvolupant determinades característiques que milloraven la seva capacitat de supervivència, les quals van convertir als primers en excel·lents màquines de matar, mentre que els segons milloraven cada vegada més les seves capacitats de detecció de perills potencials, camuflatge i fugida. Els nostres avantpassats, per contra, estaven mancats d’armes naturals, com ullals i urpes, i de la força i la velocitat que la major part de depredadors han anat adquirint gradualment en el curs de l’evolució. Per obtenir aliments, depenien de la cooperació i comunicació amb els seus congèneres, de la creació d’estris, part dels quals podien fer la funció de les armes naturals de les que els homínids no tenien, i d’una conducta flexible amb una major dependència de la que qualsevol altre animal havia tingut fins llavors, de la informació adquirida al llarg de la vida i, per tant, de la memòria, la imaginació, la previsió i la planificació.
Els nostres avantpassats van incrementar o van estabilitzar l’energia destinada al cervell augmentant la qualitat de la dieta amb un increment en el consum de carn i moll de l’os, i la cocció dels aliments, aprovisionant a femelles reproductives i la seva descendència dependent mitjançant la cria en grup i reduint l’efecte de les fluctuacions ambientals mitjançant mecanismes cognitius. D’altra banda, el nostre cervell es va beneficiar de l’energia “alliberada” per la reducció dels costos de locomoció mitjançant l’adquisició del bipedalisme i la reducció dels costos de producció mitjançant una desacceleració del nostre ritme de vida. Aquest últim factor, combinat amb la cria en grup, hauria augmentat la taxa de natalitat a la nostra espècie, molt més elevada que en altres espècies de primats properes a nosaltres.
Alguns autors creuen que la caça ha estat una de les principals pressions selectives que van promoure l’evolució del cervell i ment humans.
1ª hipòtesi
La caça requereix el potencial intel·lectual necessari per a la interacció social i la interpretació de la conducta de les preses potencials. L’adquisició de les tècniques de caça és difícil i requereix diversos anys d’educació. Va poder contribuir a l’habilitat tècnica, al desenvolupament de moviments balístics precisos en utilitzar armes llancívoles, a compartir experiències i coneixements i a enfortir els llaços d’unió entre membres del mateix grup. Nombrosos investigadors han minimitzat la contribució de la caça a l’evolució cognitiva humana, moltes vegades per raons que donen la impressió de ser més ideològiques que científiques però és difícil creure que no jugués cap paper en aquesta, encara que fos en forma indirecta (aprovisionament de carn , cooperació tecnologia).
2ª hipòtesi
La fabricació d’instruments lítics va poder propiciar l’augment de lateralització hemisfèrica que s’observa en el cervell humà, així com la utilització preferent de la mà dreta, controlada per l’hemisferi esquerre, per a realitzar manipulacions precises. La producció d’instruments implica unes elaborades capacitats d’imaginació, planificació i memòria, així com l’execució de moviments seqüencials que porten a la consecució de l’útil imaginat prèviament. La dependència dels útils va poder promoure la selecció de gens relacionats amb l’augment de capacitat de seqüenciació motora, planificació i imaginació. Aquestes últimes capacitats estan relacionades amb el còrtex prefrontal lateral.
La seqüenciació motora depèn de diverses regions cerebrals, entre les que es troben els ganglis basals, el cerebel i diverses regions del còrtex cerebral esquerre, com parts del còrtex prefrontal i premotor i àrees corticals de les que depenen la producció i comprensió del llenguatge. En moltes d’aquestes regions es produeix l’expressió del gen FOXP2, relacionat amb l’habilitat lingüística. Basant-se en aquestes dades, el neurobiòleg William Calvin va proposar la hipòtesi que la construcció d’instruments i la realització de moviments balístics precisos van poder haver promogut l’evolució del llenguatge, constituint un exemple del que Darwin deia «canvi funcional amb continuïtat anatòmica» [37]. Diverses dades provinents de la neuroanatomia, la neuroimatge i la genètica semblen donar suport a la hipòtesi de Calvin: si durant un llarg període de temps no haguéssim estat perfeccionant l’habilitat per tallar i utilitzar eines, potser mai hauríem arribat a desenvolupar el llenguatge.
3ª hipòtesi
Tot i aquestes dades, alguns científics resten importància a la influència de la tecnologia en l’evolució cerebral i cognitiva humana, i sostenen que la principal pressió selectiva que va contribuir a aquesta evolució va ser el progressiu increment de la complexitat social. La interacció amb altres membres del grup i la necessitat de compartir informació amb ells i influir en la seva conducta, van poder constituir pressions selectives que van promoure el desenvolupament del llenguatge. Van començar a adquirir importància les capacitats de detectar que altres éssers humans posseeixen estats mentals propis, la necessitat d’inferir quins eren aquests estats mentals i predir, en conseqüència, les seves possibles conductes futures [38]. A aquestes capacitats se les coneix com Teoria de la Ment i semblen ser una especialització pròpia de l’ésser humà, encara que els grans simis manifesten els precursors essencials d’aquesta capacitat, si bé en forma rudimentària.
Segons Alexander [39], la competició amb els membres del propi grup per diversos recursos, i el perill que representaven els membres de grups estranys, va iniciar una nova «carrera d’armaments» la qual va produir una coevolució similar a la que té lloc entre caçadors i preses, que va afavorir les capacitats cognitives i, possiblement tècniques. La intel·ligència social, la Teoria de la Ment, la capacitat d’inferir les intencions dels altres, la planificació i la flexibilitat conductual es van poder veure reforçades per aquesta competició intraespecífica que potser expliqui, en part, alguns dels aspectes més foscos de l’ésser humà. Els exemples de canibalisme purament alimentari, perpetrats amb tota probabilitat per membres aliens al propi grup, que es donen al llarg del Plistocè en diferents espècies d’homínids, i l’elevada taxa d’homicidi, inter i intragrupal, entre els actuals caçadors-recol·lectors, donen suport a la hipòtesi d’Alexander [40].
- Entorn social, flexibilitat conductual i plasticitat cerebral
El que tots els investigadors sí que semblen donar suport és que pràcticament no hi ha aprenentatge si no hi ha entorn social i que l’increment d’aquell està en funció directa de l’increment d’aquest. De fet, l’explicació d’això podria estar en certa particularitat que presenten tots els mamífers, especialment intensificada en el cas humà: que gran part del seu desenvolupament es porta a terme mentre l’individu ja està interactuant amb l’entorn, ja que hi ha un llarg període de desenvolupament extrauterí entre el seu naixement i el seu total desenvolupament o maduresa.
Aquest període és molt llarg en els primats i excepcional en l’ésser humà, únic gènere que sembla presentar una fase de «infantesa» en la seva ontogènia. En el cas humà, el període que dista entre la infància i la pubertat s’ha fet extremadament longeu, i d’això en diem infantesa. Aquest període, com el de la infància, és extraordinàriament ric en estímuls per a l’individu; tots dos junts suposen, per tant, una «guany» exponencial en el potencial d’aprenentatge. Per tant, com més llarg sigui el període de desenvolupament extrauterí en un àmbit social, més s’incrementarà el potencial de l’aprenentatge de l’individu. El que la complexitat cognitiva estigui relacionada amb el temps de desenvolupament ontogenètic i amb el desenvolupament cerebral sembla demostrar-se pel fet que el cervell dels nens superdotats creix durant més temps, sent, a més, un cervell més emmotllable o flexible que el dels altres nens [41].
El desavantatge adaptatiu d’aquest llarg període d’immaduresa és que requereix d’una constant cura i alimentació, però l’avantatge és que permet a l’organisme en desenvolupament la incorporació d’una gran quantitat de coneixements, el desenvolupament de la seva cognició, sintonitzada amb el medi ambient social i natural circumdant.
Una vida llarga implica enfrontar-se a un major nombre de situacions imprevistes i experimentar un major nombre de riscos i fluctuacions ambientals, el que requereix al seu torn una major dependència de l’aprenentatge per sobreviure. La llarga infància dels membres del gènere Homo va fer que aquest aprenentatge tingués lloc mentre els joves es trobaven sota la protecció familiar. El període que coneixem com adolescència va permetre un allargament addicional del període de desenvolupament cerebral i més temps per a l’aprenentatge de les habilitats necessàries per a un adult. L’adolescència sembla haver aparegut relativament tard en l’evolució humana: potser va aparèixer durant l’últim milió d’anys.
¿Som diferents gràcies al llenguatge?
Per què actualment som l’única espècie humana del planeta ?, què va succeir amb les altres espècies del gènere Homo?, quina diferència qualitativa ens ha permès ser els últims de la genealogia humana? Potser aquesta última pregunta és clau, perquè la majoria dels especialistes estan d’acord en que la major part de les diferències entre Homo sapiens i els nostres avantpassats no són qualitatives, sinó quantitatives. Un exemple de les nostres habilitats cognitives que no pot faltar en aquest capítol és el llenguatge.
Durant anys, els paleoantropòlegs experts en el tema del llenguatge han buscat la pedra filosofal que pogués donar compte de les capacitats lingüístiques de les espècies de la geneaologia humana. Des del punt de vista anatòmic sobre els òrgans que permeten emetre sons complexos, la clau podia estar tant en la posició de la laringe com en la flexió de la base del crani. Phillip Lieberman, Jeffrey Laitman i Ignacio Martínez, per citar tres dels millors experts en aquests temes, han investigat la morfologia del crani de diverses espècies del gènere Homo, sense obtenir un clar consens en les seves observacions. Per als dos primers investigadors, l’espècie Homo habilis tenia una laringe en posició elevada, com succeeix en els simis antropoïdeus i en els nostres nens i nenes menors de dos anys. En conseqüència, ni Homo habilis ni per descomptat els hominins anteriors (per exemple, els australopitecs i els paràntrops) tindrien capacitat per a la parla. Pensen que ni tan sols els neandertals tindrien les mateixes capacitats que nosaltres per emetre un llenguatge raonablement articulat. Ignacio Martinez no opina el mateix, ja que les seves reconstruccions de cranis d’Homo habilis (OH 24) i Homo ergaster (KNM ER 3733) troben valors en l’angle de la flexió de la base del crani significativament més elevats que en els australopitecs o en els simis antropodeus.
La forma de l’os hioide dels neandertals, com el trobat a l’esquelet del jaciment de Kebara (Israel), dóna suport també a la hipòtesi que aquests humans tinguessin una notable capacitat per al llenguatge.
En aquest debat, la genètica també ha aportat dades molt rellevants. Des de finals del segle passat se sap que el gen FoxP2 està implicat en el llenguatge humà, i s’expressa en el còrtex i altres zones del cervell durant l’embriogènesi. No és un gen específic del nostre llinatge (s’ha estudiat molt en aus). El que resulta interessant és que mitjançant seqüenciació de DNA s’ha pogut determinar que els neandertals posseïen la mateixa variant d’aquest gen que els sapiens actuals. Per aquest motiu, cal pensar que els neandertals probablement tenien capacitat per desenvolupar un llenguatge, el que no implica que la utilitzessin.
Des de fa temps, tots els experts saben que al neocòrtex ha dos regions ben definides relacionades amb el llenguatge. L’àrea de Broca es localitza a la part posterior de l’hemisferi esquerre (tercera circumvolució frontal), pròxim a zones relacionades amb activitats motores. El paper de l’àrea de Broca està relacionat amb l’acció de parlar i construir les frases amb les que ens expressem. L’àrea de Wernicke es localitza a la part posterior de l’hemisferi esquerre (circumvolució temporal), en una regió associativa del neocòrtex pròxima a regions amb capacitat per a l’audició. El paper de l’àrea de Wernicke es relaciona amb la nostra capacitat per escoltar i comprendre (descodificar) el que se’ns diu.
Si bé tots els experts reconeixen el paper de les àrees de Broca i Wernicke, ara se sap que en realitat la nostra capacitat per al llenguatge resideix en tot el cervell. Aquesta hipòtesi té la seva explicació en la complexitat de les diferents llengües que parlem els humans actuals. Es podria dir que el llenguatge té diferents nivells de dificultat. Els ximpanzés, que també posseeixen les seves corresponents àrees de Broca i Wernicke, són capaços d’aprendre un cert repertori de paraules «humanes», que no poden pronunciar, però que s’entenen a la perfecció. Els ximpanzés poden aprendre el llenguatge dels sordmuts, i entendre no només unes 150 paraules, sinó les combinacions de diverses d’elles; és a dir, comprenen frases més o menys complicades. Si els ximpanzés poden comunicar-se amb nosaltres a través d’un llenguatge humà, què podríem dir de les altres espècies del gènere Homo? Potser la resposta estigui en acceptar que en cada pas de l’evolució de la nostra genealogia hem anat pujant esglaons d’una llarga escala, que ha conduït de moment al màxim nivell possible d’expressió i comprensió del llenguatge.
Gairebé cap paleoantropòleg nega la possibilitat que les espècies del gènere Homo hagin tingut llenguatges rudimentaris, amb capacitat per a la comunicació més enllà dels sons que poden emetre els simis antropoïdeus en el seu medi. Al cap i a la fi, la nostra vida es va desenvolupar en espais oberts, en els quals cridar servia de poc. Podem, tanmateix, afirmar que l’enorme complexitat de les paraules que utilitzem precisa del concurs de tota l’escorça cerebral. Amb les àrees de Broca i Wernicke ja no n’hi ha prou, perquè les paraules poden tenir significats implícits de moviment, so, gust, plaer, por, dolor, etc., o referir-se a conceptes abstractes i intangibles, com l’amor, el futur o la llibertat.
A manera de conclusió
On resideix la diferència entre els nostres avantpassats africans de fa 200.000 anys i els actuals habitants del planeta? Per a diversos investigadors, aquesta diferència rau en mutacions d’uns pocs gens reguladors. Aquests gens haurien tingut una selecció positiva i s’haurien estès molt ràpidament en les poblacions humanes. Aquestes mutacions haurien estat les responsables del nostre enorme progrés tecnològic, sense oblidar per descomptat la socialització de l’art ancestral del Paleolític. Amb una única mutació gènica podem assolir resultats espectaculars i arribar a fenotips completament diferents, depenent del canvi operat en el gen regulador. Les diferències genètiques quantitatives entre els ximpanzés i la nostra espècie són certament petites. Tot just superen el 1,5% del genoma. No obstant això, la funció de cada un dels gens que ens separen d’ells pot tenir conseqüències qualitatives de gran envergadura.
Entre els gens identificats fins al moment com a possibles responsables de la mida cerebral destaquen el gen ASP M i el gen MCPH 1, que actuarien sobre la capacitat de proliferació de les cèl·lules immadures precursores de la cèl·lules nervioses (neurones i cèl·lules glials). Aquests gens reguladors semblen haver tingut (i continuen tenint) una forta pressió selectiva cap a l’increment de les variants més freqüents en la població mundial. Altres gens, com l’NRCAM, intervenen en la regulació de les connexions entre les neurones i tindrien, per tant, un paper crucial en la complexitat cerebral. Beatriz Cubelos i el seu equip han identificat dos gens homeobox, Cux1 i Cux2, que actuen en la connexió de les dendrites de les capes II i III del còrtex cerebral. És possible que la selecció natural hagi actuat sobre certes variants d’aquests i altres gens, que ens han procurat un cervell més eficaç en les seves funcions cognitives, com la memòria operativa i l’autoconsciència.
Sens dubte no som exactament iguals als primers representants d’Homo sapiens del Plistocè Mitjà. Llevat d’excepcions ben conegudes, com la del celacant (Latimeria chalumnae), que ha romàs inalterat vivint en aigües abissals des de fa gairebé 400 milions d’anys, les espècies no són estàtiques. Les espècies continentals estan exposades a canvis continus del medi. La nostra espècie amb prou feines porta vivint al planeta uns 200.000 anys, però hem estat exposats a tots els medis permesos pel nostre metabolisme i la nostra bioquímica, incloses les altures elevades, les àrees desèrtiques, el fred glacial o els tòrrids calors tropicals. A més, la cultura ha actuat com un potent factor ambiental, que (per bé o per mal) nosaltres mateixos generem. No pot estranyar, per tant, que el nostre genoma estigui fixant variants adequades a la nostra peculiar forma de vida.
Podem preguntar-nos si el que anomenem de manera genèrica intel·ligència està relacionat únicament amb mutacions específiques en el nostre genoma. És evident que tenim una gran memòria operativa, que ens permet comunicar-nos amb un llenguatge complex. Tenim una teoria de la ment molt desenvolupada i practiquem diferents habilitats artístiques. Ens preguntem pel nostre origen i per l’origen dels bacteris o de les estrelles. Però si qualsevol de nosaltres, que ens considerem tan àmpliament intel·ligents, quedéssim en la més completa solitud en un medi rural (i no diguem enmig del bosc del que provenim) a la mercè de les nostres habilitats per sobreviure probablement seriem incapaços d’aconseguir aliment. Si el lloc en qüestió es troba en l’hemisferi nord i el nostre aïllament esdevingués a la tardor o a l’hivern, probablement moriríem de fred en molt poc temps. Llavors ens sorgeixen moltes preguntes: on queden els nostres grans avantatges evolutius respecte a un neandertal o un Homo erectus?, de què ens servirien els canvis genètics suposadament fixats en el nostre genoma?, qui es podria comunicar amb nosaltres en aquest cas hipotètic?, de què ens serviria saber interpretar una partitura de Mozart o escriure un bell poema?, podríem preparar un plat d’alta cuina o aplicar coneixements d’enginyeria? Tots sabem que aquests coneixements serien inútils en aquestes condicions. Potser sobreviurien els individus entrenats per a això, però no succeiria el mateix amb la immensa majoria de nosaltres. La nostra evolució ha seguit el seu propi camí cap a una socialització molt desenvolupada. Sempre hem estat primats socials, però ara ho som en grau extrem. Depenem els uns dels altres, perquè hem construït un món a la mida de les necessitats d’una societat cada vegada més complexa i formada per grups o «professions» altament especialitzades. Estem adaptats a un medi social molt exigent. La nostra espècie ha fet un salt gegantí cap a la complexitat social: en això resideix el nostre èxit, però també el major perill.
Les societats «primitives» que encara queden a la Terra han avançat fins a un mode de vida que encaixa en la cultura que anomenem «el Neolític». Aquesta cultura va sorgir en diversos punts del planeta fa entre 10.000 i 5.000 anys, als inicis del període Holocè. Diverses poblacions van desenvolupar una nova forma d’obtenir recursos per a la seva subsistència, mitjançant la domesticació d’animals salvatges i el cultiu sistemàtic de plantes comestibles. Aquest canvi econòmic va substituir en gran mesura a la caça i la recol·lecció de productes silvestres. Finalment, la «revolució neolítica» es va estendre com una taca d’oli i va canviar la forma de vida de la immensa majoria dels habitants del planeta. El Neolític va aparèixer al sud-oest, sud-est i nord d’Àsia, al Sàhara i al cinturó del Sahel, a Nova Guinea, en regions d’Amèrica Central i en regions andines de l’Amèrica del Sud. La revolució neolítica va comportar un creixement demogràfic molt significatiu, sens dubte influït més per l’increment de la natalitat que pel descens de la mortalitat. Aquest creixement demogràfic va suposar moviments de població cap a la conquesta de nous territoris, necessaris per alimentar una població que a poc a poc va començar a créixer de manera exponencial. Tot això en detriment dels caçadors i recol·lectors, que buscaven el seu aliment pels territoris conquistats. La revolució neolítica va comportar la construcció d’habitatges i la dels primers pobles, va introduir la producció de ceràmica, necessària per conservar els excedents de les collites, la pedra polida i un ric repertori de nous útils relacionats amb la recol·lecció de les plantes cultivades i la guarda dels animals domesticats. El Neolític ha estat clau en la distribució actual de les diferents llengües i les seves variants, així com en la fixació de determinades mutacions genètiques en les poblacions humanes.
Pel que fa als europeus, és important assenyalar la regió coneguda com «Creixent Fèrtil», localitzada al sud-oest d’Àsia. Aquí va sorgir un dels centres més influents del Neolític. Aquesta zona geogràfica té forma de mitja lluna i comprèn des de l’extrem del golf Pèrsic fins a l’extrem sud del Corredor Llevantí, passant pel nord de l’actual estat de Síria. A causa de la seva posició geogràfica estratègica, la influència d’aquesta regió es va deixar notar a Europa, l’Iran, el nord de l’Índia i el nord d’Àfrica. El cultiu de cereals i la domesticació de cabres i ovelles van arribar aviat a totes aquestes regions. A més, la influència de la revolució neolítica del Creixent Fèrtil es va notar en la difusió de tres famílies lingüístiques: indoeuropees, afroasiàtiques i dravídiques, de les que deriven un gran part dels idiomes que coneixem avui en dia, des de l’extrem més occidental d’Europa fins a l’Índia, incloent el nord d’Àfrica i la península d’Aràbia. Malgrat aquest continu progrés, no està de més comentar que, actualment, els pobles del tercer món sobreviuen en pitjors condicions que els grups humans del Neolític. A més, aquelles poblacions que han patit o pateixen guerres poden tornar a formes de vida similars a les del Neolític amb preocupant facilitat. De fet, salvant alguns matisos, una bona part de la població espanyola dels anys 1940-1960 va viure en condicions molt similars a les dels pobles neolítics de fa 7.000 anys. No estem, per tant, tan lluny del nostre passat.
Entre les poblacions actuals que encara no han arribat a la revolució neolítica comptem amb els pigmeus de la regió del Congo, a l’Àfrica Equatorial, els Hadza de Tanzània o els Ache de Paraguai. Tots ells viuen de la caça i la recol·lecció, encara que ja s’han contaminat del progrés del que anomenem «la civilització». El genoma dels components de tots aquests pobles és com el dels altres humans del planeta i, tanmateix, no han assolit el grau de complexitat cultural que caracteritza les civilitzacions més avançades. Així que hi ha d’haver alguna cosa més que mutacions genètiques perquè la nostra espècie hagi arribat a cotes tecnològiques impensables fa tan sols un parell de centenars d’anys. És evident que la immensa majoria dels nostres avenços tecnològics han estat potenciats per la gran sociabilitat d’Homo sapiens, que ha crescut exponencialment durant els darrers anys gràcies als avenços en microelectrònica i telecomunicacions. La suma de moltes ments i l’especialització han estat claus en el progrés tecnològic. Els éssers humans som en l’actualitat totalment interdependents. Cada un de nosaltres desenvolupa un paper complementari amb el dels altres membres de la societat. Formem així una veritable xarxa social, dins i fora d’internet, en la qual les nostres neurones formen un «cervell col·lectiu», que substitueix a la individualitat. És molt possible que en les societats primitives hi hagués individus amb una alta capacitat creativa. No obstant això, les innovacions protagonitzades per aquests individus desapareixien en molt poc temps sense arribar més enllà de, com a molt, uns quants centenars de quilòmetres. A més, l’escassa longevitat impedia que la immensa majoria dels individus arribessin a aquesta edat en què atresorem l’experiència que lleguem a les següents generacions. Si la complexitat del llenguatge està relacionada amb la mutació d’algun gen regulador, és obvi que aquesta mutació només hauria estat el catalitzador d’una major capacitat per a la comunicació, la suma de solucions i l’aparició d’idees emergents (innovacions). A més, els humans actuals ens recolzem des de fa relativament poc temps en els pensaments, reflexions i fets que van deixar escrits altres humans que ens van precedir. El primer deure d’un científic, per exemple, consisteix a beure de les fonts escrites, de les reflexions i els resultats anteriors. Aquestes fonts, sumades a les seves pròpies observacions, seran el punt de partida de les seves investigacions i determinaran un possible avanç del coneixement.
En resum, a les possibles mutacions que han acabat fixant-se per selecció positiva en el genoma de les actuals poblacions del planeta hem d’afegir-hi la connexió virtual entre els centenars o milers d’individus que formem cada població, i la que globalment formen totes les poblacions del planeta. No cal que les nostres neurones entrin en contacte directe per desenvolupar aquest cervell col·lectiu. Tots coneixem que la tecnologia ja ha prescindit de la connectivitat física per transmetre informació. Els éssers humans vam fer el mateix fa relativament poc temps i estem donant un pas transcendental cap al futur, potser d’una nova espècie.
Lectures de referència
Agustí, Jordi; Bufill, Enric i Mosquera, Marina (2012). El precio de la inteligencia. Barcelona. Crítica. (Col. Drakontos). 256 pàgs.
Allman, John Morgan (2003). El cerebro en evolución. Barcelona. Ariel. (Col. Ariel neurociencia). 1ª impr. 240 pàgs.
Arsuaga, Juan Luis i Martínez, Ignacio (2006). La especie elegida : la larga marcha de la evolución humana. Madrid. Ed. Temas de Hoy. 1ª impr. 448 pàgs.
Briones, Carlos; Fernández, Alberto i Bermúdez, José María (2015). Orígenes. El universo, la vida, los humanos. Barcelona. Crítica. Pàgs. 341-469
Campàs, Joan (2014): 18. Els primers Homo: Homo habilis. Curs en línia: «Els orígens de l’art i l’evolució humana: l’Homo significans». En línia a http://cv.uoc.edu/adf/~04_999_01_u07/llico_18.pdf
Campàs, Joan (2014): 19. El poblament d’Euràsia: l’Homo ergaster/erectus. Curs en línia: «Els Orígens de l’art i evolució humana: l’homo significans». En línia a http://cv.uoc.edu/adf/~04_999_01_u07/llico_19.pdf
Campàs, Joan (2014): 20. Homo antecessor, rhodesiensis, heidelbergensis i floresiensis. Curs en línia: «Els orígens de l’art i evolució humana: l’homo significans». En línia a http://cv.uoc.edu/adf/~04_999_01_u07/llico_20.pdf
Campàs, Joan (2014). 21. L’Homo neanderthalensis. Curs en línia: «Els orígens de l’art i evolució humana: l’homo significans». En línia a http://cv.uoc.edu/adf/~04_999_01_u07/llico_21.pdf
Campàs, Joan (2014). 22. L’Homo sapiens. Curs en línia: «Els orígens de l’art i evolució humana: l’homo significans». En línia a http://cv.uoc.edu/adf/~04_999_01_u07/llico_22.pdf
Campàs, Joan (2014). 31. Sobre els orígens del llenguatge. Curs en línia: «Els orígens de l’art i evolució humana: l’homo significans». En línia a http://cv.uoc.edu/adf/~04_999_01_u07/Llico_31.pdf
Carbonell i Roura, Eudal (2005). Homínidos : las primeras ocupaciones de los continentes. Barcelona. Ariel. (Col. Ariel prehistoria ). 864 pàgs.
Cela-Conde, Camilo i Ayala, Francisco (2011). Senderos de la evolución humana. Madrid. Alianza ed. (Col. Alianza ensayo,188). 7ª impr. 632 pàgs.
Leakey, Richard Erskine (2000). El origen de la humanidad. Barcelona. Debate. (Col. Temas de Debate). 1ª impr. 224 pàgs.
Striedter, Georg F. (2005). Principles of brain evolution. Sunderland MA. Sinauer Associates. 436 pàgs.
Tattersall, Ian (2012). Los señores de la tierra. La búsqueda de nuestros orïgenes humanos. Barcelona. Pasado y Presente. 1ª impr. 352 pàgs.
Notes
[1] Paola Villa i Wil Roebroeks: «Neandertal demise: An archaeological analysis of the modern superiority complex». PLoS ONE 9: e96424. En línia a http://journals.plos.org/plosone/article?id=10.1371/journal.pone.0096424
[2] Günter Bräuer i Richard Leakey: «The ES-11693 Cranium from Eliye Springs, West Turkana, Kenya». Journal of Human Evolution (1986) 15, 289-312. En línia a http://in-africa.org/wp-content/uploads/2012/12/Brauer-Leakey-1986-JHE-Eliye-Springs.pdf
[3] Rebecca l. Cann, Mark Stoneking i Allan C. Wilson: «Mitochondrial and human evolution». Nature 325, 31 – 36 (01 January 1987)
[4] Ewen Callaway: «Humans may have trekked across the Arabian peninsula 125,000 years ago, on their way to Asia». Nature, 27 January 2011. En línia a http://www.nature.com/news/2011/110127/full/news.2011.55.html
[5] Arsuaga, Juan Luis, i Martínez, Ignacio (2006). La especie elegida. La larga marcha de la evolución humana. Madrid. Temas de Hoy. 1ª impr. 448 pàgs.
[6] Ruff, Chistopher .B., Thinkaus, Erik, Holliday, Trenton W. (1997). «Body mass and encephalization in pleistocene Homo». Nature 387: 173-176. En línia a https://www.researchgate.net/publication/14074734_Body_mass_and_encephalization_in_Pleistocene_Homo_Nature
[7] Catherine Gianaro: «Lahn’s analysis of genes indicates human brain continues to evolve». The University of Chicago Chronicle. September 22, 2005, Vol. 25 No. 1. En línia a http://chronicle.uchicago.edu/050922/brainevolution.shtml
[8] Mekel-Babrov, Nitzan, Gilbert, Sandra L., Evans, Eric J. Vallender, Jeffrey R. Anderson, Richard R. Hudson, Sarah A. Tishkoff, Bruce T. Lahn (2005). «Ongoing adaptive evolution of ASPM, a brain size determinant in Homo sapiens». Science 309: 1720-22. En línia a https://www.researchgate.net/publication/7611130_Ongoing_Adaptive_Evolution_of_ASPM_a_Brain_Size_Determinant_in_Homo_Sapiens
Patrick D. Evans, Sandra L. Gilbert, Nitzan Mekel-Bobrov, Eric J. Vallender, Jeffrey R. Anderson, Leila M. Vaez-Azizi, Sarah A. Tishkoff, Richard R. Hudson, Bruce T. Lahn (2005). «Microcephalin, a gene regulating brain size, continues to evolve adaptively in humans». Science 309: 1717-1720. En línia a http://www.evolocus.com/Publications/Evans2005.pdf
[9] Aboitiz, F. (1994). «The evolution of brain and organization in vertebrales: a program for research». Biological Research. 27: 15-27
[10] P. Brown, T. Sutikna, M. J. Morwood, R. P. Soejono, Jatmiko, E. Wayhu Saptomo i Rokus Awe Due (2004): «A new small-bodied hominin from the Late Plestocene of Flores, Indonesia». Nature 431: 1055-1061. En línia a http://www.readcube.com/articles/10.1038/nature02999
[11] Leslie C. Aiello i Wheeler, P.: «Brains and guts in human and primate evolution: the expensive organ hypothesis». Current Anthropologie 36, 199-221. Veure en línia a http://www.scielo.br/scielo.php?script=sci_arttext&pid=S0100-84551997000100023
[12] Gunz, Phillipp, i Neubauer, Simon, Maureille, Bruno, i Hublin, Jena-Jacques (2010): «Brain development after birth differs between Neanderthals and modern humans». Current Biology 20, R921-R922. En línia a http://www.sciencedirect.com/science/article/pii/S0960982210012820
[13] Daniel E. Lieberman i Robert C. McCarthy (1999): «The ontogeny of cranial base angulation in humans and chimpanzees and its implications for reconstructing pharyngeal dimensions». Journal of Human Evolution 36, 487-517. En línia a http://www.people.fas.harvard.edu/~skeleton/pdfs/1999c.pdf
[14] Dunsworth, H.M., Warrener, A.G., Deacon, T., Ellison, P.T., i Pontzer, H. (2012): «Metabolic hypothesis for human altriciality». Proceedings of the National Academy of Sciences, USA 103, 18178-18183.
[15] Daniel J. Miller , Tetyana Duka , Cheryl D. Stimpson , Steven J. Schapiro , Wallace B. Baze i altres («Prolonged myelination in human neocortical evolution». Proceedings of tje National Academy of Sciences, USA 109, 16480-16485. En línia a http://www.pnas.org/content/109/41/16480.full.pdf
[16] Bookstein, F., Schafer, K., Prossinger, H., Seidler H, Fieder M, Stringer C, Weber GW, Arsuaga JL, Slice DE, Rohlf FJ, Recheis W, Mariam AJ, Marcus LF. (1999): «Comparing frontal cranial profil·les in archaic and modern Homo by morphometry analysis». The Anatomical Record. 15;257(6):217-24.
[17] Brumner, Emiliano (2004). «Geometric morphometrics and paleoneurology: Brain shape evolution in the genus Homo». Journal of Human Evolution, 47: 279-303. En línia a http://www.sciencedirect.com/science/article/pii/S0047248404001149
Brumner, Emiliano i Ralph L. Holloway (2010): «A bivariate approach to the widening of the frontal lobes in the genus Homo». Journal of Human Evolution, 58: 138-146. En línia a http://www.sciencedirect.com/science/article/pii/S0047248409002206
[18] Weaver, Anne H. (2005). «Reciprocal evolutionm of the cerebellum and neocòrtex in fòssil humans». Proceedings of the National Academy of Sciences of the USA 102: 3576-80. En línia a http://www.pnas.org/content/102/10/3576.full
[19] Rilling, James K., i Insel, Thomas.R. (1999). «The primate neocòrtex in comparative perspective using magnètic resonance imaging». Journal of Human Evolution, 37: 191-223. En línia a http://ibrarian.net/navon/paper/The_primate_neocortex_in_comparative_perspective_.pdf?paperid=6078862
[20] Preuss, T.D. (2004): «What is it like to be a human?», a Gazzaniga Michael S. (ed.). The cognitive neurosciences III. Cambridge MA. MIT Press. Pàgs. 5-22
[21] Rilling, J.K. (2008): «Neuroscientific approaches and Applications within anthropology». American Journal of Physical Anthropology 51: 2-32.
[22] Semendeferi, Katerina, Damasio, Hanna, Frank, Randall i Hoesen, Gary W. van (1997). «The evolution of the frontal lobes: a volumètric analysis based on three-dimensional reconstructions of magnètic resonance imatge scans of human and ape brains». Journal of Human Evolution, 32: 375-388.
[23] Katerina Semendeferi, Este Armstrong, Axel Schleicher, Karl Zilles, and Gary W. Van Hoesen (2001): «Prefrontal còrtex in humans and apes: a comparative study of area 10». American Journal of Physical Anthropology, 144: 224-241. En línia a http://cs.brown.edu/people/tld/projects/cortex/course/suggested_reading_list/supplements/documents/SemendeferietalAJPA-01.pdf
[24] Coqueugniot H., Hublin J.-J., Veillon F., Houët F., Jacob T., (2004). «Early brain growth in Homo erectus and implications for cognitive ability». Nature, 431 : 299-302. En línia a http://www.eva.mpg.de/evolution/staff/hublin/pdf/Coqueugniot_et_al_04_Nature.pdf
[25] Parker, S.T., i Mc Kinney, M.L. (1999). Origins of intelligence. The evolution of cognitive development in monkeys, apes and humans. Baltimore. The John Hopkins University Press. 424 pàgs.
[26] Bogin, B. (2006): «Modern human life history: the evolution of human childhood and fertility», a Paine R.R., Hawkes, K. (eds.). The evolution of the human life history. Santa Fe, Nuevo México. School of American Research Press. Pàgs. 197-230.
[27] Allman, John Morgan (1999). Evonving brains. Nova York. Scientific American Library.224 pàgs.
[28] Kristen Hawkes, James F. O’Connell, Nicholas G. Blurton Jones, H. Alvarez, i E. L. Charnov (1998): «Grandmothering, menopause and the evolution of human life histories». Proceedings of the National Academy of Sciences of the USA, 95: 1336-1339.
[29] Klein, R.G. (1999). The human career. Human biological and cultural origins. Chicago. The University of Chicago Press. 840 pàgs.
[30] Caspari, Rachel, i Lee, Sang-Hee (2004): «Older age becomes common late in human evolution». Proceedings of the National Academy of Sciences of the USA, 101: 10895-900. En línia a http://www.pnas.org/content/101/30/10895.full
[31] Pollard, Katherine S., Salanna, Sophie R., Lambert, Nelle, Marie-Alexandra Lambot, Sandra Coppens, Jakob S. Pedersen, Sol Katzman, Bryan King, Courtney Onodera, Adam Siepel, Andrew D. Kern, Colette Dehay, Haller Igel, Manuel Ares, Jr, Pierre Vanderhaeghen and David Haussler (2006): «An RNA gene expressed during cortical devewlopment evolved rapidly in humans». Nature 443: 167-172.
[32] Herz, Joachim, Chen, Ying (2006): «Reelin, lipoprotein receptors and synaptic plasticity». Nature Reviews Neuroscience 7, 850-859 (November 2006). En línia a http://www.nature.com/nrn/journal/v7/n11/full/nrn2009.html
[33] Martínez-Cerdeño Verónica, Galazo Maria José, Cavada Carmen i Clascá, Francisco (2002): «Reelin immunoreactivity in the adult primate brain: intracellular localization in projecting and local circuit neurons of the cerebral còrtex, hippocampus and the subcortical regions». Cerebral Cortex 12: 1298-1311.
[34] Cáceres, M., Suwyn, C., Maddox, M., Thomas, J.W., Preuss, T.M. (2007): «Increased cortical expression of two synaptogenic thrombospondins in human brain evolution». Cerebral Cortex. 17: 2312-21.
[35] Finlayson, C. (2004). Cognitive capacity and its outward expression: Pleistocen homínids across a space-time mosaic of opportunity. Gibraltar. Museo de Gibraltar, pàgs. 1-18.
[36] Aiello, Leslie C. i Wheeler, Peter (1995). «The expensive tissue hypothesis: the brain and digestive System in human and primate evolution». Current Anthropology, Vol. 36, No. 2 (Apr., 1995), pp. 199-221. En línia a https://www.jstor.org/stable/2744104?seq=1#page_scan_tab_contents
[37] Calvin, William H. (2004). A brief history of the mind. Oxford. Oxford University Press. 240 pàgs.
[38] Cosmides, Leda, i Tooby, John (2000): «The evolution of adaptations for decoupling and metarepresentation», a Sperber, Dan (ed.). Metarepresentations. Nova York. Oxford University Press. 460 pàgs. En línia a https://monoskop.org/images/9/9c/Sperber_Dan_(ed.)_Metarepresentations_A_Multidisciplinary_perspective_2000.pdf
[39] Alexander, R. (1990). How did humans evolve? Reflections on the uniquely unique species. Michigan. Ann Arbor. Museum of Zoology (Special publication nº 1). The University of Michigan. En línia a http://qcpages.qc.edu/Biology/LahtiSites/RDAlexander/Pubs/Alexander90.pdf
[40] Ghiglieri, Michael Patrick (2005). El lado oscuro del hombre : los orígenes de la violencia masculina. Barcelona. Tusquets. (Col. Metatemas, 86). 1ª impr. 384 pàgs.
[41] P. Shaw, D. Greenstein, J. Lerch, L. Clasen, R. Lenroot, N. Gogtay, A. Evans, J. Rapoport and J. Giedd (2006): «Intellectual ability and cortical development in children and adolescents». Nature 440: 676-679.En línia a https://www.semanticscholar.org/paper/Intellectual-ability-and-cortical-development-in-Shaw-Greenstein/48357eaa6dd8e31536316d7006c32e6a66e36ccb/pdf
Dr. Joan Campàs Aura digital
Estudis d’Arts i Humanitats de la UOC
Curs Som les nostres connexions
Material en format pdf
Tema 0. Presentació i justificació
Tema 1. El cervell ens enganya
Tema 2. El cervell també enganya l’autopercepció del cos
Tema 3. Anatomia cerebral bàsica
Tema 4. Mite: Només utilitzem el 10 % del nostre cervell
Tema 5. Mite: L’homosexualitat (l’hetero, la bi, la trans) és una elecció
Tema 6. Mite: Els fills hereten la intel·ligència dels pares
Tema 7. Mite: Internet i els videojocs són addictius
Tema 8. Mite: El cervell és un ordinador
Tema 9. La memòria
Tema 10. Com definir el «jo»? El cervell i la «joïtat»
Tema 11. L’evolució del cervell (1)
Tema 12. L’evolució del cervell (2)
Tema 13. L’evolució del cervell (3)
Tema 14. L’evolució del cervell (4)
Tema 15. Cervell humà i evolució